
Journal of Clinical Images and Medical Case Reports
ISSN 2766-7820
Short Commentary - Open Access, Volume 4
Paradigm shift: Bacterial quorum sensing and possible control targets
Eric O Omwenga1*; Silas O Awour2; Francisco M Goycoolea3
1Kisii University, School of Health Sciences, Kisii, Kenya.
2Microbiology Department, Jaramogi Oginga Odinga Teaching and Referral Hospital, Kisumu, Kenya.
3School of Food Science and Nutrition, University of Leeds, Leeds, LS2 9JT, United Kingdom.
*Corresponding Author : Eric O Omwenga
Kisii University, School of Health Sciences, Kisii, Kenya.
Email: omorieric@kisiiuniversity.com
Received : Sep 02, 2023
Accepted : Sep 29, 2023
Published : Oct 06, 2023
Archived : www.jcimcr.org
Copyright : © Omwenga EO ( 2023).
Abstract
QS Designates a cell-to-cell communication process that enables bacteria to collectively modify their behaviour in response to changes in the cell density and species composition of the surrounding microbial community. These processes involve the production, release and group-wide detection of extracellular signalling molecules, which are generically called Autoinducers (AIs). It controls various genes which are responsible for various phenotypes like bioluminescence, the secretion of virulence factors, and the formation of biofilms in bacteria. Quorum quenching inhibits QS and the substances that inhibit it are called quorum sensing inhibitors. Several chemical compounds and enzymes mediate inhibition of QS, such as, lactonases, acylases and oxidoreductases. Other than these, there are some non-enzymatic methods for quorum quenching and also some plant phytochemicals have been found to inhibit it. Blocking of QS by QS Inhibition (QSI) may play an important role to disrupt biofilm formation in a device associated infection and chronic drug resistant infection. More researches are required in this area related with QS and QSI. However, some chemicals have been found to be mimicking the quorum sensing AIs activities like serotonin and rosmaric acid.
Keywords: Quorum sensing; Quorum quenching; Autoinducers; Quorum sensing inhibitors.
Citation: Omwenga EO, Awour SO, Goycoolea FM. Paradigm shift: Bacterial quorum sensing and possible control targets. J Clin Images Med Case Rep. 2023; 4(10): 2632.
Bacterial quorum sensing and its autoinducers
The last two decades or so we have witnessed a remarkable change in the understanding of how bacteria live and function. In this regard, it is now firmly established that bacteria have evolved several mechanisms to facilitate the interaction among them. One such strategy is the ability to coordinate gene expression in response to population density, a process known as Quorum Sensing (QS) [1-4]. QS Designates a cell-to-cell communication process that enables bacteria to collectively modify their behaviour in response to changes in the cell density and species composition of the surrounding microbial community [3,4]. These processes involve the production, release and group-wide detection of extracellular signalling molecules, which are generically called Autoinducers (AIs). AIs are produced at basal levels and their concentration increases with growth. Because the signals can diffuse through membranes, their concentration inside cells approximates the concentration in the environment. Upon reaching a critical concentration, the signal molecules can bind to and activate receptors inside bacterial cells. These receptors can then alter gene expression to activate behaviours that are beneficial under the particular condition encountered. As this phenomenon occurs in a cell-density-dependent manner, it has been termed hence the term quorum sensing [3,5]. Processes that are controlled by QS, such as bioluminescence, the secretion of virulence factors, and the formation of biofilms, are unproductive and costly when undertaken by a single bacterial cell, but become effective when undertaken by the group [4]. Alternative hypotheses to the original QS proposal have also been advanced. One of them is based in the fact that in addition to cell density, the concentration of AIs in the natural environment is determined by many other biotic and abiotic factors, such as the spatial distribution of cells and the diffusional characteristics of the environment [6-10], this was called Diffusion Sensing (DS). Yet another concept of Efficiency Sensing (ES) was introduced in an attempt to unify both QS and DS hypotheses [10]. In recently published studies [11,12], a single intracellular protein governs the critical transition from an individual to a coordinated population response during quorum sensing: Origins of primordial language. Vila-Sanjurjo, et al., [13], further proposed that bacterial communication mediated by QS is analogous to a critical phenomenon that can be explained by physical laws that govern phase transitions in nature, such as the conversion of water into vapour by temperature. Formal theoretical treatments, namely the Ising model [11] and percolation theory [11,12] have been applied to QS phenomena. These recent approaches, although do not contradict the basic concepts of QS, they account for several aspects that were not hitherto considered in the original QS hypothesis, such as the underlying reason of the existence of a critical threshold in the synchronous response, and its network character at the genetic level.
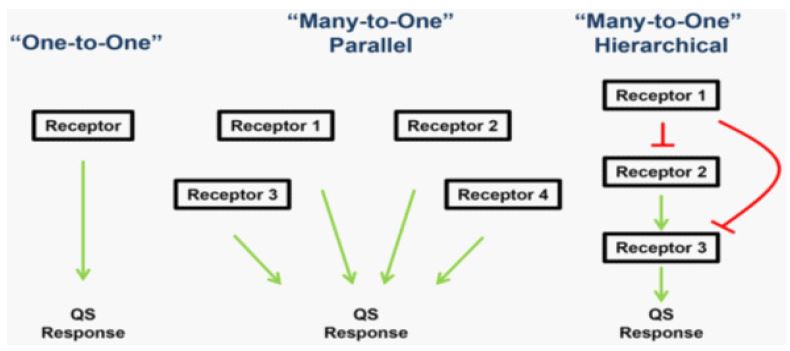
QS is mediated by diffusible signal molecules that reflect the density and physiological state of their population. Many pathogenic bacteria, such as Staphylococcus aureus, Vibrio cholera or Pseudomonas aeruginosa etc., regulate the expression of their virulence factors by QS. Such organisms use various networks or circuits to undertake their QS as shown in Figure 1 below [14]. Many classes of AIs have been described to date (Figure 2) [14].
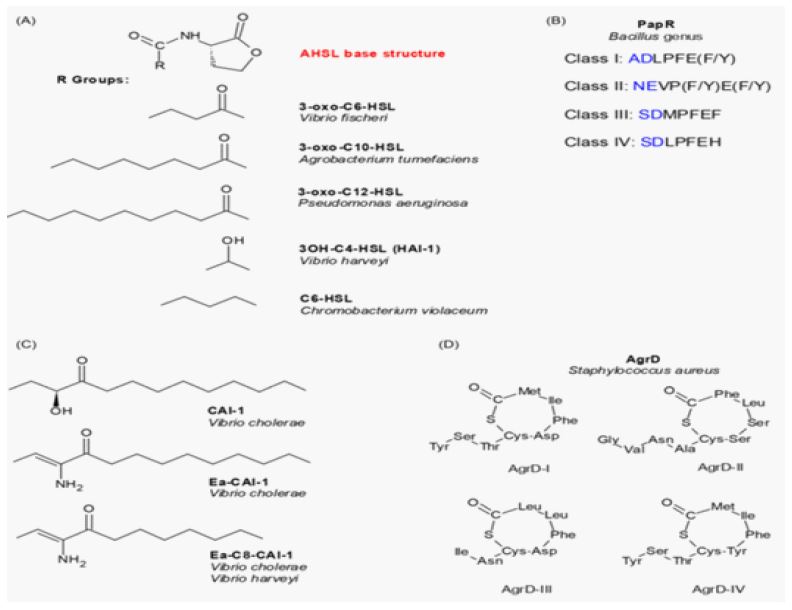
The most intensely studied AIs are the N-Acyl Homoserine Lactones (AHLs) of Gram-negative bacteria, the peptides of Gram-positive bacteria and a class of AIs termed AI-2, whose structures remain unknown in most cases [5]. AHLs are usually detected through binding to and activation of cytoplasmic receptor proteins, which dimerize upon signal detection and can bind to promoter regions of target genes to activate or repress their transcription [15]. Peptides are usually detected through binding to membrane sensor proteins of the two-component system family, although some can also be transported to the cytoplasm before interacting with their receptors [16,17]. On the other hand, AI-2 binds a periplasmic protein and then interacts with either a two component system or a transporter depending on the organism [18,19]. Binding to a membrane-associated sensor kinase causes the activation of a phosphorelay cascade, which results in the activation or repression of a response regulator, culminating in altered gene expression [16,18]. It is now widely recognized that many bacterial species utilize QS as part of their regulatory machinery [3,5,20]. Of interest, we now know that bacterial virulence is in many cases controlled by QS [5]. This has led to a burst in research and the role of QS in the virulence of multiple human pathogens has been studied in molecular detail.
Selected quorum sensing circuits
Staphylococcus aureus as a model for gram positive bacteria
Many Gram-positive bacteria utilize peptide QS systems to control gene expression and S. aureus (Figure 3) has served as a model to study bacterial peptide signalling [16]. S. aureus is also a very dangerous opportunistic pathogen which has been increasingly associated with antibiotic resistance [21]. It is a bacterium that is a member of the human microbiota and is found in approximately 30% of the adult population. It possesses a lot of virulence traits/factors and can display very rapid transmission, aiding its importance as a very dangerous human pathogen [22]. In addition this bacterium is known to form biofilms on many surfaces, including indwelling devices such as urethral stents/catheters hence posing a serious risk for infections related to this bacterium some of which are too hard to control [15].

The peptide-based quorum system contributes immensely to the virulence traits of S. aureus and it is encoded by the accessory gene regulator (agr) locus [16]. The agr system is thought to regulate over 70 genes, 23 of which are known virulence factors [21]. Of the virulence factors regulated by agr, there are two classes: The first class contains virulence factors involved in attachment to the host and immune evasion, while the second class contains genes involved in the production of exoproteins associated with invasion and toxin production. It has been thought that the activation of the agr system essentially switches the bacterium from an adhesive, colonizing commensal to an invasive and aggressive pathogen [15].
Pseudomonas aeruginosa as a model for gram negative bacteria
In a large number of Gram-negative bacteria, QS is mediated via N-acyl-homoserine lactones (AHLs) [23]. AHL signalling molecules are composed of a homoserine lactone (HSL) ring with an acyl chain; the acyl-chain length typically varies from C4 to C18 and may be modified, most often by a 3-oxo substituent, a 3-hydroxy substituent, or by its degree of unsaturation. Pseudomonas aeruginosa is an example of Gram-negative bacterium which is an opportunistic pathogen that is commonly associated with nosocomial and severely burned individuals’ infections, and is a leading cause of death in severe respiratory infections, such as chronic lung infections in patients with cystic fibrosis [24]. Infections with P. aeruginosa are difficult to eradicate, due to their high levels of antibiotic resistance and growth in biofilms [25].
At least three intertwined QS systems and one orphan autoinducer receptor affect the ability of P. aeruginosa to cause disease (Figure 4). Two of these systems, Las and Rhl, rely on the production of N-Acyl Homoserine Lactones (AHLs) as the signalling molecules (AIs) [15]. In the Las system, N-3-oxododecanoyl-homoserine lactone (3OC12-HSL) is produced by the enzyme encoded by the las I gene. When P. aeruginosa reaches a certain threshold density, 3OC12-HSL binds to the transcriptional activator Las R. Las R, in turn, dimerizes and binds to target promoters to control gene expression [15]. Similarly, in the Rhl system, the rhlI gene encodes the enzyme involved in the production of N-butyryl-homoserine lactone (C4-HSL). As with 3OC12-HSL, C4-HSL binds to its cognate transcriptional regulator, Rhl R, to control the activity of target promoters [26]. The rhl system is controlled by the Las system at both transcriptional and post-transcriptional levels. Besides LasR and RhlR, P. aeruginosa encodes an orphan receptor protein, QscR, which can sense 3OC12- HSL to control its own regulon [15,23].
Quorum sensing regulates the production of several extracellular virulence factors, promotes biofilm maturation and regulates the expression of antibiotic efflux pumps, meaning that it has a key role in the pathogenesis of P. aeruginosa [15,27]. The Las and Rhl systems regulate the timing and production of multiple virulence factors, including elastase, alkaline protease, exotoxin a, rhamnolipids, pyocyanin, lectins and superoxidase dismutase. The expression of these two quorum sensing systems has also been linked to the regulation of biofilm formation [15].
Other than virulence traits regulations some of the AIs have also been found to have negative effects on the host immune system. For instance, 3OC12-HSL induces Interleukin (IL)-8 secretions from human bronchial epithelial cells and induces COX-2 production [28]. It also inhibits lymphocyte proliferation, down regulates the production of tumour necrosis factor alpha (TNF-a) and IL-12, and activates T cells to produce gamma-interferon [29]. 3OC12-HSL also induces apoptosis in macrophages and neutrophils, suggesting that this molecule not only enhances the expression of virulence genes but also affects immune responses to infection [28,30]. Additionally, 3OC12-HSL has been shown to directly affect one of the major regulators of immune responses, NFkB [31].

These amongst many models clearly indicate that bacteria use AIs to develop various virulence traits through QS cascades that assist them survive in the host hence inducing an infection and resistance to antibiotics through such mechanisms like biofilm formation. Therefore, a new generation of drugs is needed to quench/inhibit the activity of bacterial autoinducers and the entire QS circuit hence managing them and the conditions they can cause.
Antiquorum sensing strategies
Introduction
Inhibiting bacterial QS could be an alternative strategy towards eradication or prophylaxis of bacterial-borne infections. This is because targeting and blocking QS circuits of microorganisms may disarm their collective virulent traits, hence, making them susceptible to elimination by host immune system or to low doses of antibiotics. Anti-virulent agents from either natural products or synthetic ones possessing QS inhibitory activity might not pose selective pressure on pathogens for survival. Hence, they are likely to attenuate the rapid emergence of drug resistance [32,33]. Disruption of QS-regulated processes has been accepted to provide two advantages: i) reduced accumulation of virulence factors at the infection site, and ii) the collective power of pathogens being dismantled. These strategies are collectively known as Quorum Quenching (QQ) [33].
There are several quorum quenching strategies available through which the process of quorum sensing can be interrupted. They can be classified broadly into three major groups, namely: i) Inhibition of auto inducers like AHL synthesis by blocking the synthase proteins like Lux I-type; ii) Enzymatic destruction of autoinducers like AHLs molecules by AHL-acylase and AHL-lactonase that will prevent them from accumulating; and iii) Interference with signal receptors or blockage of formation of AHL/Lux R (autoinducer - receptor) complex [35,36]. These strategies are summarized in Figure 5.
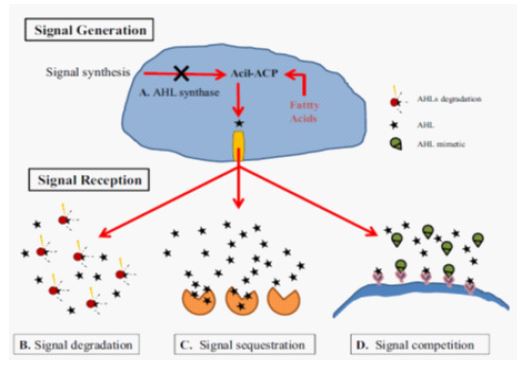
Quorum quenching enzymes
Among the three strategies some work has been done on enzymatic degradation of autoinducers by the enzymes called quorum quenching (QQ) enzymes. Such group of enzymes were first isolated from Bacillus species [37] and they have been classified into three major groups based on their mode of action against the AHL autoinducer. Lactonases forms one of the classes of QQ enzymes and their mode of action is mainly by opening the homoserine (HSL) ring [38,39]. Another class of QQ enzymes are the acylases that mainly cleave the acyl side chain from the HSL ring [36,39]. Oxidoreductase QQ enzymes is another group that is known to catalyse the oxidation or reduction of acyl side chain [36].
The QQ enzymes are mainly derived from bacteria, fungi, plants and legumes. For instance, bacterial AHL acylases reported includes AiiD from Ralstonia sp. XJ12B [40], AhlM from Streptomyces sp. [41], PvdQ and QuiP from P. aeruginosa PAO1 [42] and AiiC from Anabaena sp. PCC7120 [42]. The acylases HacA and HacC produced by P. syringae B728a have shown to degrade quorum sensing signal AHLs [43]. On the other hand, a good number of AHL lactonases have also been reported and among them includes AttM from A. tumefaciens c58 [44], AiiA from Bacillus sp. 240B1 [37], AiiB from A. tumefaciens [37], QIcA from Acidobacteria sp. [45] and AidH from Ochrobactrum sp. T63 [46]. Also, legumes like alfalfa, clover, lotus, peas and yam bean have been reported to contain AHL degrading enzyme lactonase [36,47]. Altogether, these, amongst many more, constitute a library of QQ enzymes that have been discovered and could be of great help towards managing QS. Perhaps, once more knowledge is gained on how to improve their stability, administration and release they may become a group of antivirulence therapeutics. A lot of ongoing research is also focused on sequestration and signal completion at the receptor level. Several reports have been made of antagonists that have been reported and some examples are discussed here below.
Medicinal plants phytochemicals as antagonists of bacterial quorum sensing
Plants are reservoir of an extremely rich library of secondary metabolites like tannins, terpenoids, alkaloids, polyphenols, flavonoids, alkaloids, lectins, polypeptides, polyacetylenes, essential oils, amongst many more, which have been found in vitro to have antimicrobial properties against both Gram-positive and Gram-negative bacteria [48]. Plants use these compounds in various ways. For example, flavonoids are the key pigments that reproduce via biotic pollination; they are also involved in UV-light filtration and symbiotic nitrogen fixation; and as chemical messengers, they act as physiological regulators, and cell-cycle inhibitors. Essential oils, on the other hand, are most commonly associated with the distinctive flavours and aromas of many plants and are quite abundant in nature [49]. Modes of activity of some of these plant compounds have been known (Figure 6) while in others it’s still not clear on how they act against the microbial cells [48].
Medicinal plants extracts and their phytochemicals have been found to provide a most promising source for searching of natural QS inhibitory compounds. New compounds mined from algae, fungi, bacteria and higher plants have been discovered to possess antiquorum sensing and antibiofilm formation properties. Such natural compounds, have been found to possess such advantages like imparting less selective pressure on the microorganisms of interest due to their low biocidal activity. Thus, the chances of development of resistance are in principle minimized. This is mainly because they are structurally similar to the cognate quorum sensing autoinducer signal molecules. Thus, they can easily work as antagonists of LuxR/LasR signal receptors [50,51].

For instance, furanones mined from Australian red alga, Delisea pulcha have been reported to possess Antiquorum Sensing (AQS) activity and antibiofilm formation against P. aeruginosa, V. harveyi and E. coli. It mainly inhibits the networks that are activated by the various types of AHL. They are thought to act by interfering with AHL signal from its reporter protein [52,53]. Lowery et al. [54], further indicates that halogenated furanones destabilize and accelerate Lux R turn over hence impairing the binding of the AHL to DNA to initiate transcription. On the other hand, furanones have also been found to inhibit AI-2 based quorum sensing system as they are structural mimics of lactones and tetrahydrofuran rings of quorum sensing system36. Defoirdt et al. [55]; further indicates that (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone, a natural compound produced by marine alga has the ability to inhibit swarming motility and biofilm formation in E. coli at non-lethal concentrations to planktonic growth.
The flavonoids like kaempferol, naringenin, quercetin, flavan-3-ol catechin and apigenein have been reported as inhibitors of AHL and AI-2-mediated pathogenic traits of P. aeruginosa PAO1 and E. coli O157:H7 [56,57]. They have been found to inhibit various virulence traits like biofilm formation, pyocyanin production, and elastase production, without inhibiting bacterial growth [56]. Also, reduction in the expression of several quorum sensing-controlled genes (e.g., LasI, LasR, RhlI, RhlR, LasA, LasB, phzA1 and RhlA) has also been reported in P. aeruginosa PAO1. and C. violaceum CV026 respectively with these compounds [58]. By use of structure-activity relationship (SAR) analysis, Paczkowski et al. [59] did show that the novel flavonoids like quercetin, baicalein and others which possesses dihydroxyl moieties in the flavone A-ring backbone, have been found to bind to the QS receptors, LasR and RhlR allosterically. This ends up-reducing significantly their ability to bind to DNA encoding QS regulated promoters. These findings clearly do demonstrate that phytochemicals can be potential inhibitors of bacterial QS and may form a novel class of agents that can assist mankind in dealing with conditions caused by “superbugs’’.
Many more compounds do possess these properties and have been reported. For example, furocoumarin has been found to possess QS-mediated biofilm formation in E. coli and swarming motility in P. aeruginosa PAO1 [60,61]. Limonoids from sour orange seeds such as isolimonic acid, ichangin and deacetyl nomilinic acid 17 β-D-glucopyranoside, were found to inhibit AI-2 mediated quorum sensing in V. Harveyi [62].
Also, metabolic isolates from fungi have been studied and they do possess antiquorum sensing properties. Penicilic acid and patulin produced by Penicillium radicicola and Penicillium coprobium respectively have been found to possess the QS inhibitory activity [50]. The DNA microarray-based transcriptomics showed that penicilic acid and patulin target, respectively, 60% and 45% quorum sensing genes in P. aeruginosa, thus suggesting the RhlR and LasR quorum sensing regulators [36,63].
These well-contrasted activities of these compounds, amongst many more, clearly indicate that natural QS inhibition can offer a potential solution to the multidrug resistance associated with traditional common antibiotics or antimicrobial compounds for specific bacterial pathogens. Recent comprehensive reviews on natural compounds that antagonize QS have been published [36,63]. However, further investigations are required to continue identifying potential QSIs present in natural sources, as well as elucidating the mechanism they use to antagonize QS, and the ways in which their activity may be improved. Antivirulence agents could be key in management of the emerging and re-emerging microbial infections as they will be targeting their virulence traits and not killing the microbe thus effectively exerting less selective pressure and likely reducing the current rising trend of antimicrobial resistance. One of such methods that can be used to improve drug delivery is using of nanomaterials as vehicles of transporting QSIs to target sites.
Other QQ strategies
Other strategies like non- enzymatic strategies are being explored as novel strategies in management of bacterial quorum sensing. For instance, several research groups have explored usage of monoclonal antibodies to quench AHLs - a good example is the administration of conjugated bovine serum albumin with 3oC12HSL intranasally unto mice and it did challenge Pseudomonas aeruginosa by sequestering the autoinducers while not having any impact to its viability [64-66]. Other strategies could be usage of antibodies that can bind with the chemical signals of quorum sensing molecules hence inhibiting their activity.
Quorum sensing agonists
It has also been realised that some chemicals other than the autoinducers triggers bacterial quorum sensing. Such mimickers of autoinducers are naturally produced by the human body system to assist in various physiological processes while others are phytochemicals produced by plants. For instance, Knecht et al. [67] did report that the gut derived serotonin a neurotransmitter that is synthesized by gut enterochromaffin cells does mimic autoinducer of the Las R circuit of the P. aeruginosa. This gut-derived serotonin has also been reported to be responsible for regulation of functions such as bone development, immune responses, gut motility, and platelet aggregation [68]. From the studies carried out by Knecht et al. [67] it was clearly demonstrated that gut derived serotonin activates the Las quorum sensing pathway, which leads to greater infectivity of P. aeruginosa. It was also deduced that serotonin increased significantly the virulence traits production of elastase and protease in both PAO1 and JP2 cells. Biofilm formation, the mediator of on-set of pathogenicity in the hosts, was also evaluated & was deduced that JP2 cells only formed biofilms when seroserotonin, QSM or both were present. PAO1 cells also demonstrated increased biofilm formation in the presence of serotonin [64]. This form of “social cheating” in the human body perhaps could be an evolutionary characteristic for opportunistic bacteria P. aeruginosa to enhance its virulence production. On the other hand, such findings could undoubtedly be used to harness more knowledge on the complexity involved in relationships between the host and its’ microbiome. It may lead to better understanding of regulation of the microbiome by the host during infection which may play an important role in the management of GIT bacterial-related illnesses or disorders of the digestive system that are triggered by bacteria.
Other than in humans it has also been reported that some plant phytochemicals do mimic bacterial QS autoinducers. Corral-Lugo et al. [69] did report that rosmaric acid phytochemical exhibited a significantly greater activity on the RhlR QS circuit of P. aeruginosa compared to C4-HSL autoinducer on responding to equal concentrations on the ligand that was compared to the control (absence of ligand). Further work on treating the ligand with half-equimolar C4-HSL or rosmaric acid to RhlR concentrations, transcriptional activity was less than at equimolar concentrations of ligand to regulator, a clear indication of dose dependence of the response. In vivo studies also did indicate significant increase of β-Galactosidase measurements which is as a result of gene expression of P. aeruginosa in the presence of C4-HSL or rosmaric acid. Further work of rosmaric acid did indicate elevated increase in production of virulence traits like biofilm formation, pyocyanin production, and elastase synthesis with no antimicrobial activity on the dosage applied. Since rosmaric acid is a phytochemical produced by plants as a defence strategy [70], it has been postulated that such phytochemicals may decrease pathogenicity by stimulating premature expression of genes encoding proteins involved in QS-controlled functions [71,72]. This is due to the fact that the pathogen involved in this study affects both plants and animals [73]. Some phenolic compounds on the other hand have been found to elevate biofilm formation but however they have not been reported to mimic QS autoinducers [74]. These amongst some more phytochemicals could be having an agonistic activity against QS of bacteria [75-79].
It’s now clear that bacteria-to-plant and plant-to-bacteria signalling are also based on QS systems. Proteomics studies have shown that some autoinducers like HSLs modulate a number of genes expressions in legumes like Medicago truncatula [80]. Also, transcriptomic studies revealed that C6-HSL, a bacterial QS molecule produced in the rhizosphere, changed gene expression in Arabidopsis thaliana [81]. HSL signalling processes are, in part, responsible for the induced systemic resistance of plants toward bacterial pathogens [82]; these processes also modulate plant growth [83]. This clearly demonstrates that there could be an evolution with diverse signalling systems that permits mutual plant-microorganism sensing hence coexistence.
Conclusion
Quorum sensing is responsible for various virulence traits observed in bacteria and therefore forms one of the key areas that can be explored as means of eradicating antimicrobial resistances observed in the world today. QS is mediated by chemical signals (autoinducers) which are produced by pathogens in synchrony and therefore targeting these signals by either inhibiting their productivity, degradation, sequestering or having competitors for their target receptors can be a key strategy in management of most conditions produced by such groups of bacteria which have been proved to be resistant to most antibiotics. This is generally because less evolutionary pressure will be mounted unto the bacteria as such targets will not end up killing the bacteria but later inhibit a specific pathway (QS) that is involved in virulence production. Several natural compounds have been mined like flavonoids from plants and have been observed to inhibit such a pathway. Also, it has been realised that some compounds like serotonin and rosmaric acid act as agonists of quorum sensing a finding that demonstrates that signalling systems are diverse in nature most probably used as a way of co-existence. It is also clear that some of the natural compounds that inhibit quorum sensing also have a big impact on biofilm formation that is the end product of quorum sensing.
References
- Whitehead NA, Barnard AM, Slater H, Simpson NJ, Salmond JP. Quorum sensing in Gram-negative bacteria. FEMS. Microbiol. Rev. 2001; 25: 365-404.
- Waters CM, Bassler BL. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005; 21: 319-346.
- Bassler BL, Losick R. Bacterially speaking. Cell. 2006; 125: 237-246.
- Papenfort K, Bassler BL. Quorum sensing signal-response systems in Gram-negative bacteria. Nature Reviews Microbiology. 2016; 14: 576-588.
- Antunes LC, Ferreira RB. Intercellular communication in bacteria. Crit. Rev. Microbiol. 2009; 35: 69-80.
- Microbienne E, Moulin BJ. Cell-cell signalling in bacteria: Not simply a matter of quorum. FEMS Microbiol Ecol. 2009; 70: 1-19.
- Redfield RJ. Is quorum sensing a side effect of diffusion sensing? Trends Microbiol. 2002; 10: 365-370.
- Decho AW, Norman RS, Visscher PT. Quorum sensing in natural environments: emerging views from microbial mats. Trends Microbiol. 2010; 18: 73-80.
- West SA, Winzer K, Gardner A, Diggle SP. Quorum sensing and the confusion about diffusion. Trends Microbiol. 2012; 20: 586-594.
- Hense BA, Kuttler C, Müller J, Rothballer M. Does efficiency sensing unify diffusion and quorum sensing? Nat. Rev. Microbiol. 2007; 5: 230-239.
- Weber M, Buceta J. The cellular Ising model: A framework for phase transitions in multicellular environments. J. R. Soc. Interface. 2016; 13.
- Vila-Sanjurjo C, et al. Percolation theory can explain primordial forms of cell-to-cell communication. Submitted.
- Vila-Sanjurjo C. et al. A single intracellular protein governs the critical transition from an individual to a coordinated population response during quorum sensing: Origins of primordial language. 2016.
- Hawver LA, Jung SA, Ng WL. Specificity and complexity in bacterial quorum-sensing systems. FEMS Microbiology Reviews, fuw0. 2016; 14: 1-15.
- Caetano LMA, Rosana BRF, Michelle MCB, Brett BF. Quorum sensing in bacterial virulence, Microbiology. 2010; 156: 2271-2282.
- Novick RP, Geisinger E. Quorum sensing in Staphylococci. Annu.Rev. Genet. 2008; 42: 541-564.
- Pottathil M, Lazazzera BA. The extracellular Ph peptide- Rap phosphatase signalling circuit of Bacillus subtilis. Front. Biosci. 2003; 8: d32-d45.
- Ng WL, Bassler BL. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009; 43: 197-222.
- Taga ME, Semmelhack JL, Bassler BL. The LuxS dependent autoinducer AI-2 controls the expression of an ABC transporter that functions in AI-2 uptake in Salmonella typhimurium. Mol. Microbiol. 2001; 42: 777-793.
- Smith RS, Harris SG, Phipps R, Iglewski B. The Pseudomonas aeruginosa quorum-sensing molecule N-(3-oxododecanoyl) homoserine lactone contributes to virulence and induces inflammation in vivo. J. Bacteriol. 2002; 184: 1132-1139.
- George EA, Muir TW. Molecular mechanisms of agr quorum sensing in virulent staphylococci. Chem. Bio-Chem. 2007; 8: 847-855.
- Massey RC, Horsburgh MJ, Lina G, Hook M, Recker M. The evolution and maintenance of virulence in Staphylococcus aureus: A role for host-to-host transmission. Nat. Rev. Microbiol. 2006; 4: 953- 958.
- Fuqua, C. The QscR quorum-sensing regulon of Pseudomonas aeruginosa: an orphan claims its identity. J. Bacteriol. 2006; 188: 3169-3171.
- Bendiak GN, Ratjen F. The approach to Pseudomonas aeruginosa in cystic fibrosis. Semin. Respir. Crit. Care Med. 2009; 30: 587-595.
- Driscoll JA, Brody SL, Kollef MH. The epidemiology, pathogenesis and treatment of Pseudomonas aeruginosa infections. Drugs. 2007; 67: 351-368.
- De Kievit TR, Iglewski BH. Bacterial quorum sensing in pathogenic relationships. Infect. Immun. 2000; 68: 4839-4849.
- Dekimpe V, De´ ziel E. Revisiting the quorum-sensing hierarchy in Pseudomonas aeruginosa: The transcriptional regulator RhlR regulates LasR-specific factors. Microbiology. 2009; 155: 712-723.
- Smith RS, Iglewski BHP. Aeruginosa quorum-sensing systems and virulence. Curr. Opin. Microbiol. 2003; 6: 56-60.
- Skindersoe ME, Zeuthen LH, Brix S, Fink LN, Lazenby J, et al. & other authors Pseudomonas aeruginosa quorum-sensing signal molecules interfere with dendritic cell-induced T-cell proliferation. FEMS Immunol. Med. Microbiol. 2009; 55: 335-345.
- Tateda K, Ishii Y, Horikawa M, Matsumoto T, Miyairi S, et al. The Pseudomonas aeruginosa autoinducer N-3-oxododecanoyl homoserine lactone accelerates apoptosis in macrophages and neutrophils. Infect. Immun. 2003; 71: 5785-5793.
- Kravchenko VV, Kaufmann GF, Mathison JC, Scott DA, Katz AZ, et al. Modulation of gene expression via disruption of NF-kB signalling by a bacterial small molecule. Science. 2008; 321: 259- 263.
- Vattem DA, Mihalik K, Crixell SH, McLean RJC. Dietary phytochemicals as quorum sensing inhibitors. Fitoterapia. 2007; 78: 302-10.
- Kumar L, Chhibber S, Kumar R, Kumar M, Harjai K. Zingerone silences quorum sensing and attenuates virulence of Pseudomonas aeruginosa. Fitoterapia. 2015; 102: 84-95.
- Dong YH, Wang LH, Zhang LH. Quorum-quenching microbial infections: Mechanisms and implications. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2007; 362: 1201-1211.
- Truchado P, Larrosa M, Castro-Ibáñez I, Tomás-Barberán FA, Allende A. Plant food extracts and phytochemicals: Their role as Quorum Sensing Inhibitors, Trends in Food Science & Technology. 2015; 43: 189-204.
- Lade H, Paul D, Kweon JH. Quorum quenching mediated approaches for control of membrane biofouling. Int. J. Biol. Sci. 2014; 10: 550-565.
- Dong YH, Xu JL, Li XZ, Zhang LH. AIIA. An enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Prac Natt Acad Sci. U.S.A. 2000; 97: 3526-3531.
- Czajkowski R, Jafra S. Quenching of acyl-homoserine lactone-dependent quorum sensing by enzymatic disruption of signal molecules. Acta. Biochemica. Polonica. 2009; 56: 1-16.
- Leadbetter JR, Greenberg EP. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J. Bacteriol. 2000; 182: 6921-6926.
- Lin YH, Xu JL, Hu JY, Wang LH, Ong SL, et al. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol. Microbiol. 2003; 47: 849-860.
- Park SY, Kang HO, Jang HS, Lee JK, Koo BT, et al. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005; 71: 2632-2641.
- Huang JJ, Petersen A, Whiteley M, Leadbetter JR. Identification of QuiP, the product of gene PA1032, as the second acyl-homoserine lactone acylase of Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2006; 72: 1190-1197.
- Shepherd RW, Lindow SE. Two dissimilar N-acyl-homoserine lactone acylases of Pseudomonas syringae influence colony and biofilm morphology. Appl. Environ. Microbiol. 2009; 75: 45-53.
- Zhang HB, Wang LH, Zhang LH. Genetic control of quorum-sensing signal turnover in Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA. 2002; 99: 4638-4643.
- Riaz K, Elmerich C, Moreira D, Faure DA. Metagenomic analysis of soil bacteria extends the diversity of quorum-quenching lactonases. Environ. Microbiol. 2008; 10: 560-70.
- Mei GY, Yan XX, Turak A, Luo ZQ, Zhang LQ. Aid H, an alpha/beta-hydrolase fold family member from an Ochobactrum sp. strain, is a novel N-acylhomoserine lactonase. Appl. Environ. Microbiol. 2010; 76: 4933-4942.
- Fatima Q, Zahin M, Khan MSA, Ahmad I. Modulation of quorum sensing controlled behaviour of bacteria by growing seedling, seed and seedling extracts of leguminous plants. Ind. J. Microbiol. 2010; 50: 238-242.
- Bazaka K, Jacob MV, Chzanowski W, Ostrikov K. Anti-bacterial surfaces: Natural agents, mechanisms of action, and plasma surface modification. RSC. Adv. 2015; 5: 48739-48759.
- Lacobellis NS, Lo Cantore P, Capasso F, Senatore F. Antibacterial activity of Cuminum cyminum L. and Carum carvi L. essential oils. J. Agric. Food Chem. 2005; 12: 57-61.
- Vattem DA, Mihalik K, Crixell SH, McLean RJ. Dietary phytochemicals as quorum sensing inhibitors. Fitoterapia. 2007; 78: 302-310.
- Teplitski M, Mathesius U, Rambaugh KP. Perception and degradation of N-acyl homoserine lactone quorum sensing signals by mammalian and plant cells. Chem. Rev. 2011; 111: 100-116.
- Manefield M, de Nys R, Kumar N, Read R, Givskov M, et al. Evidence that halogenated furanones from Delisea pulcha inhibit acylated homoserine lactone (AHL)-mediated gene expression by displacing the AHL signal from its receptor protein. Microbiology. 1999; 145: 283-291.
- Manefield M, Rasmussen TB, Henzter M, Andersen JB, Steinberg P, et al. Halogenated furanones inhibit quorum sensing though accelerated LuxR turnover. Microbiology. 2002; 148: 1119-1127.
- Lowery CA, Dickerson TJ, Janda KD. Interspecies and interkingdom communication mediated by bacterial quorum sensing. Chem. Soc. Rev. 2008; 37: 1337-1346.
- Defoirdt T, Miyamoto CM, Wood TK, Meighen EA, Sorgeloos P, et al. The natural furanone (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone disrupts quorum sensing-regulated gene expression in Vibrio harveyi by decreasing the DNA-binding activity of the transcriptional regulator protein LuxR. Environ. Microbiol. 2007; 9: 2486-2495.
- Vikram A, Jayaprakasha GK, Jesudhasan PR, Jayaprakasha GK, Pillai SD, et al. Suppression of bacterial cell-cell signalling, biofilm formation and type III secretion system by citrus flavonoids. J. Appl. Microbiol. 2010; 109: 515-527.
- Vandeputte OM, Kiendrebeogo M, Rasamiravaka T, Stévigny C, Duez P, et al. The flavanone naringenin reduces the production of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology. 2011; 157: 2120-2132.
- Vandeputte OM, Kiendrebeogo M, Rajaonson S, Diallo B, Mol A, et al. Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010; 76: 243-253.
- Paczkowski JE, Mukherjee S, McCready AR, Conga J, Aquino CJ, et al. Flavonoids suppress Pseudomonas aeruginosa virulence through allosteric inhibition of quorum-sensing receptors. The Journal of Biological Chemistry. 2017; 292: 4064-4076.
- Girennavar B, Cepeda ML, Soni KA, Vikram A, Jesudhasan P, et al. Grapefruit juice and its furocoumarins inhibits autoinducer signalling and biofilm formation in bacteria. Int. J. Food Microbiol. 2008; 125: 204-208.
- Zahin M, Hasan S, Aqil F, Khan MS, Husain FM, et al. Screening of certain medicinal plants from India for their anti-quorum sensing activity. Indian J. Exp. Biol. 2010; 48: 1219-1224.
- Vikram A, Jesudhasan PR, Jayaprakasha GK, Pillai SD, Patil BS. Citrus limonoids interfere with Vibrio harveyi cell-cell signalling and biofilm formation by modulating the response regulator LuxO. Microbiology. 2011; 157: 99-110.
- Rasmussen TB, Givskov M. Quorum sensing inhibitors: a bargain of effects. Microbiology. 2006; 152: 895-904.
- Miyairi S, Tateda K, Fuse ET, Ueda C. Immunization with 3-oxo dodecanoyl - 1 - homoserine lactone - protein conjugate protects mice for lethal Pseudomonas aeruginosa lung infection. J Med Microbiol. 2006; 55: 1381 - 1387.
- Kaufmann GF, Sartorio R, Lee SH, Mee JM, Attobell LJ, et al. N- acyl homoserine lactone - mediated bacterial quorum sensing. J. Am. Chem. Soc. 2006; 128: 2802-2803
- Kaufmann GF, Park J, Mee JM, Ulevitch RJ, Janda KD. The quorum-quenching antibody RS2-1G9 protects macrophages from the cytotoxic effects of Pseudomonas aeruginosa quorum sensing signalling molecule N - 3 - Oxo - dodecanoyl homoserine lactone. Mol. Immunol. 2008; 45: 2710 - 2714.
- Knecht LD, O’Connor G, Mittal R, Liu XZ, Daftarian P, et al. Serotonin Activates Bacterial Quorum Sensing and Enhances the Virulence of Pseudomonas aeruginosa in the Host, E Bio Medicine. 2016.
- Berger M, Gray JA, Roth BL. The expanded biology of serotonin. Annu. Rev. Med. 2009; 60: 355-366.
- Corral-Lugo A, Daddaoua A, Ortega A, Espinosa-Urgel M, Krell T. Rosmarinic acid is a homoserine lactone mimic produced by plants that activates a bacterial quorum-sensing regulator. Science Signalling. 2016; 9: 409.
- Venturi V, Fuqua C. Chemical signalling between plants and plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2013; 51: 17-37.
- Walker TS, Bais HP, Déziel E, Schweizer HP, Rahme LG, et al. Pseudomonas aeruginosa-plant root interactions. Pathogenicity, biofilm formation, and root exudation. Plant Physiol. 2004; 134: 320-331.
- Cao H, Baldini RL, Rahme LG. Common mechanisms for pathogens of plants and animals. Annu. Rev. Phytopathol. 2001; 39: 259-284.
- Plyuta V, Zaitseva J, Lobakova E, Zagoskina N, Kuznetsov A, et al. Effect of plant phenolic compounds on biofilm formation by Pseudomonas aeruginosa. APMIS. 2013; 121: 1073-1081.
- Degrassi G, Devescovi G, Solis R, Steindler L, Venturi V. Oryza sativa rice plants contain molecules that activate different quorum-sensing N-acyl homoserine lactone biosensors and are sensitive to the specific AiiA lactonase. FEMS Microbiol. Lett. 2007; 269: 213-220.
- Teplitski M, Robinson JB, Bauer WD. Plants secrete substances that mimic bacterial N-acyl homoserine lactone signal activities and affect population density-dependent behaviours in associated bacteria. Mol. Plant Microbe Interact. 2000; 13: 637-648.
- Subramoni S, Gonzalez JF, Johnson A, Péchy-Tarr M, Rochat L, et al. Bacterial subfamily of LuxR regulators that respond to plant compounds. Appl. Environ. Microbiol. 2011; 77: 4579-4588.
- Gao M, Teplitski M, Robinson JB, Bauer WD. Production of substances by Medicago truncatula that affect bacterial quorum sensing. Mol. Plant Microbe Interact. 2003; 16: 827-834.
- Ferluga S, Venturi V. OryR is a LuxR-family protein involved in interkingdom signaling between pathogenic Xanthomonas oryzae pv. oryzae and rice. J. Bacteriol. 2009; 191: 890-897.
- Perez-Montano F, Jimenez-Guerrero I, Sanchez-Matamoros RC, Lopez-Baena FJ, Ollero FJ, et al. Rice and bean AHL-mimic quorum-sensing signals specifically interfere with the capacity to form biofilms by plant-associated bacteria. Res. Microbiol. 2013; 164: 749-760.
- Mathesius U, Mulders S, Gao M, Teplitski M, Caetano-Anollés G, et al. Extensive and specific responses of a eukaryote to bacterial quorum sensing signals. Proc. Natl. Acad. Sci. U.S.A. 2003; 100: 1444-1449.
- von Rad U, Klein I, Dobrev PI, Kottova J, Zazimalova E, et al. Response of Arabidopsis thaliana to N-hexanoyl-DL homoserine - lactone, a bacterial quorum sensing molecule produced in the rhizosphere. Planta. 2008; 229: 73-85.
- Schikora A, Schenk ST, Stein E, Molitor A, Zuccaro A, et al. N-acyl-homoserine lactone confers resistance toward bio trophic and hemibiotrophic pathogens via altered activation of AtMPK6. Plant Physiol. 2011; 157: 1407-1418.
- Schenk ST, Stein E, Kogel KH, Schikora A. Arabidopsis growth and defense are modulated by bacterial quorum sensing molecules. Plant Signal. Behav. 2012; 7: 178-181.