
Journal of Clinical Images and Medical Case Reports
ISSN 2766-7820
Review Article - Open Access, Volume 4
Linking the gut microbiota to brain dysfunction in bipolar disorder: Evidence from neuroimaging
Tingting Huang1,2#; Yanchao Huang3#; Yushan Shang1,2; Chunxiao Dai1,2; Qixiu Zhang1,2; Jian Xie1,2*; Haisheng Zhang1,2*
1Department of Clinical Psychology, Affiliated Hangzhou First People’s Hospital, Zhejiang University School of Medicine, Hangzhou 310006, China.
2Zhejiang Provincial Clinical Research Center for Mental Disorders, Hangzhou, China.
3Department of Clinical Medicine, Zhejiang Chinese Medical University, Hangzhou 310053, China.
#These authors have contributed equally to this work.
*Corresponding Author : Haisheng Zhang, MD
Department of Clinical Psychology, Affiliated Hangzhou First People’s Hospital, Zhejiang University School of Medicine, No.261, Huansha Road, Hangzhou 310006, China.
Ph: 86-571-56006902, Fax: 86-571-56006902;
Email: zhanghaisheng3065@126.com
Jian Xie, MM
Zhejiang University School of Medicine, No. 261, Huansha Road,Hangzhou 310006, China.
Email: srxjian@163.com
Received : Jun 13, 2023
Accepted : Jul 04, 2023
Published : Jul 11, 2023
Archived : www.jcimcr.org
Copyright : © Zhang H & Xie J (2023).
Abstract
The gut microbiota can influence the brain function through a variety of ways, but its role in the etiology of Bipolar Disorder (BD) is yet unknown. The brain and the gut are linked through a communication pathway called the Microbiota-Gut-Brain Axis (MGBA). The current review aims to summarize the gut microbiota, neuroimaging endophenotype and its correlation in BD patients. The results demonstrated that patients with BD have drastically altered microbial diversity and taxonomic composition. Affective and cognitive-related neural networks dysfunction were found in BD patients. Additionally, BD-related microorganisms were discovered to be connected to BD-typical brain network functional connectivity properties. These investigations give preliminary evidence of the intimate connection between gut microbiota and brain networks in BD, which helps to elucidate the role of the MGBA mechanism in the pathogenesis of the condition.
Keywords: Bipolar disorder; Gut microbiota; Neural networks; MGBA.
Citation: Huang T, Huang Y, Shang Y, Xie J, Zhang H, et al. Linking the gut microbiota to brain dysfunction in bipolar disorder: Evidence from neuroimaging. J Clin Images Med Case Rep. 2023; 4(7): 2493.
Introduction
Bipolar Disorder (BD) is a prevalent chronic mental illness marked by recurrent manic or hypomanic episodes that may alternate with depressed episodes. According to the World Mental Health Survey Initiative, the lifetime prevalence of BD was 2.4%, with BD-I accounting for 0.6%, BD-II accounting for 0.4%, and subliminal BD accounting for 1.4% [1]. It results in a high disease burden and severe impairment in cognitive and social functions. However, the pathogenesis of BD remains unknown due to a lack of specific or objective biomarkers.
BD is a genetically complex disorder in which both genetic and environmental factors play a role [2]. The neurobiological reasons of BD included neuroinflammation, abnormalities in neuroplasticity, and brain dysfunction. Manic and depressed BD episodes cause the activation of neuroinflammatory pathways. Microglia release proinflammatory cytokines and neurotrophic factors during acute episodes of inflammation, lowering synaptic function and preventing the adverse effects that follow.The buildup of proinflammatory cytokines in chronic inflammation alters the neural pathways involved in mood and cognition, particularly in the hippocampus, prefrontal cortex, and amygdala regions [3,4]. According to recent studies on the gut-brain axis, the gut microbiota play a critical role in orchestrating brain development and behavior, and the immune system is now recognized as a key regulator of these interactions [5]. The intestinal bacteria in the gastrointestinal tract regulate CNS function, behavior, and peripheral immunological responses, which may contribute to BD.
BD may be understood in the broader context of Microbiota-Gut-Brain Axis (MGBA) dysfunction based on the research that is currently available. The objective of the current review is to provide a brief introduction to the MGBA, focus on earlier research on changes in the gut microbiota in BD patients, describe the neuroimaging endophenotype in BD patients, and discuss structural and functional changes in the brain in brain-gut interactions. We go into further detail on the shortcomings of earlier research and provide possible avenues for further investigation.
The MGBA
The human intestinal microbiota, which is made up of 10 to 100 trillion microorganisms that live in our gastrointestinal system and have a genome at least 100 times larger than our own [6]. The second genome of humans is now understood to be the gut microbiota, which is also a crucial component of human biology and a crucial environmental determinant for human health. The gut microbiota is dynamic and complex, constantly interacting with the organs and systems of our bodies and influencing host homeostasis and health status [7]. Yet, there are still gaps in our knowledge of how the gut microbiota affects brain function, particularly in humans, where it is difficult to draw conclusions from direct experiments.
The development of the brain itself in the growing infant has been shown to be influenced by the microbiota, and the brain could affect the gut microbiota and its metabolite secretion [8]. In recent years, it has been discovered that the gut and the brain interact reciprocally, leading to the concept of the MGBA. The MGBA is a significant and intricate bidirectional communication axis that connects the brain and the gut [9]. Through neural pathways, the neuroendocrine system, and the immune system, the brain can influence the gut microbiota and the release of its metabolites; conversely, the gut microbiota may produce a variety of metabolites, including short-chain fatty acids, inflammatory factors, and neurotransmitters, which either directly or indirectly control brain function through the enteric-vagus nervous system, enteric-mucosa-brain barrier, and hypothalamic-pituitary-adrenal (HPA) axis [9] (Figure 1).
The brain can influence the intestinal microbiota directly or indirectly through host-enteric microbiota signaling or by influencing the intestinal environment [10]. The sympathetic and parasympathetic nervous systems are parts of the Autonomic Nervous System (ANS), which mediates communication between the Central Nervous System (CNS) and Enteric Nervous System (ENS). This interaction can alter the physiology of the gut (muscle, mucosa, vasculature, and immune system), which in turn alters the composition and functionality of the gut microbiota [11,12]. The brain indirectly affects the gut by influencing the mucosal immune response, affecting intestinal epithelial permeability, and altering gastrointestinal secretion. The brain can regulate a variety of signaling molecules, including catecholamines, serotonin, dynorphin, GABA, and cytokines [13].These neurotransmitters also have an impact on gut motility, nutrition absorption, gastrointestinal innate immunity, and gut microbiota [14]. The foundational stage for microbial colonization of the gut occurs during the first year of life, which is also the fastest and most dynamic period of postnatal brain development. Infant research revealed that alterations in the gut microbiome were related to the emergence of infant cognition [15]. According to the study, infants with higher levels of Bacteroides had higher cognitive scores at two years old, whereas infants with higher levels of alpha diversity had worse scores on the overall composite score, the visual reception scale, and the expressive language scale [15]. Infant neurological development and growth closely resemble the evolutionary trajectory of the infant microbiota [16,17]. Stress has been shown to impair brain growth and raise the danger of neurological and mental disorders [18]. Preclinical research showed that stress can change the composition of the gut microbiota in animal models of stress, such as mother separation [19,20]. On day 3 after separation, there were fewer Lactobacilli present; these alterations in the microbiome were linked to stress [20]. The abundance of the genera Bacteroides and Lactobacillus was shown to decrease in mice exposed to social stressors, while the abundance of the genera Clostridium rose. These changes were associated with the levels of proinflammatory cytokines [21,22].
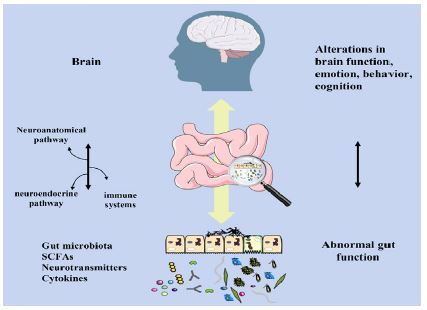
Communication from the gutmicrobiota to the CNS can occur through several different pathways. The gut microbiota regulates the maturation of the immune system, the regulation of hormones (including the HPA axis), metabolic activity, and barrier function [17]. Changes in gut microbiota and decreased Short Chain Fatty Acids (SCFA), a byproduct of gut microbiota, lead to immune cell activation, such as that of macrophages, monocytes, and microglia, which may compromise the Blood-Brain Barrier (BBB) and gut and encourage immune system activation on both the peripheral and cellular levels [23,24]. The BBB integrity and brain health were revealed to be influenced by the gut microbiota and its byproducts [25]. According to a study on animals, pathogen-free mice with a normal gut microbiota showed lower BBB permeability than germ-free mice [26]. Additionally, intestinal barrier permeability and function could be influenced by the gut microbiota [27,28], Leaky gut may be one of the underlying causes of disorders that also involve concurrent downstream BBB abnormalities [29]. The BBB acts as a semipermeable barrier to control the flow and exchange of nutrients and chemicals between the circulating blood and the brain, protecting the specific microenvironment of the CNS [30]. The BBB is crucial to brain development because it shields the growing brain from dangers outside of the body. Additionally, the gut microbiota controls brain function through the HPA axis [31,32]. The parameters of HPA axis hyperactivity have also been observed in germ-free mice in response to stress [33]. Probiotics based on Lactobacillus and Bifidobacterium have been shown to improve learning, memory, and symptoms that resemble sadness and anxiety as well as correct HPA axis dysfunction brought on by stress [34,35]. It has also been reviewed how the HPA axis mediates early life bidirectional communication between the gut microbiota and brain development [36]. According to these results, colonization of the gut microbiota is essential for controlling HPA responses and brain function. Besides, the gut microbiota regulates brain function through metabolic pathways. The bacterial metabolites SCFA and neurotransmitters including Gamma-Aminobutyric Acid (GABA), serotonin (5-HT), and dopamine, which are crucial for brain function, could be produced by the gut microbiota. Through immunological, endocrine, vagal, and other humoral pathways, SCFAs can directly or indirectly affect mood, learning, memory, and other aspects of brain function [23]. 90% of 5-HT was discovered to originate from the gut, and enteric 5-HT networks may be able to control how the adult enteric nervous system develops [37,38]. The experiment revealed that germ-free mice have an immature ENS that becomes normal after being colonized by a normal microbiota. Serotonin release was then seen to begin, followed by the activation of the 5-HT4 receptor [38]. Additionally, the hippocampus of GF mice had greater concentrations of noradrenaline, dopamine, and 5-HT [ 39]. By controlling the metabolites of tryptophan in the gut, the gut microbiota may be able to influence brain 5-HT.
Gut microbiota changes in BD patients
Recent research found that mental illnesses could also be understood in the wider context of MGBA dysfunction. Mental illnesses such as schizophrenia, Major Depressive Disorder (MDD), and BDwere shown to alterin gut microbiota traits [40-42]. In comparison to controls, BD is associated with lower levels of microbial diversity and differing relative abundances of bacterial phyla. Several investigations on humans found that BD patients have a less diverse gut microbiota [43-47]. In brief, we outlined the gut microbiome characteristics associated with mood disorders in our prior review from 2019 [48]. In these studies, it was discovered that the gut microbiota of BD patients tended to have higher levels of the phylum Actinobacteria, the order Coriobacteriales, the family Coriobacteriaceae, the genus Enterobacteriaceae, the lower genus Faecalibacterium, and the abundance of Bacteroides. It has been established that Actinobacteria, Coriobacteriales, Coriobacteriaceae, and Bacteroides are involved in the metabolism of lipids and sugars. Flavonifractor was also linked to the impact of oxidative stress and inflammatory responses [48]. Several studies have demonstrated that patients with bipolar disorder have altered gut microbial makeup. Increased abundance of phylum Actinobacteria [43,48,49] and Firmicutes [43,44,49], order Coriobacteriales [48], family Coriobacteriaceae [48], Clostridiaceae [46], Enterobacteriaceae [48], Streptococcaceae [45,47], genus Flavonifractor [48], Clostridium [43,49], Bifidobacterium [43,49], Oscillibacter [43,49], Streptococcus [43,49] were found in BD patients, while phylum Bacteroidetes [43,45,48], family Ruminococcaceae [45,47], Faecalibacterium [48] were decreased. A reduced abundance of SCFA-producing bacteria was found in the gut microbiota of BD patients [44,50]. Clinical manifestations of BD may be partly explained by variations in gut microbiota composition. Collinsella was found to be more prevalent in BD-II patients than BD-I patients, according to McIntyre et al [46]. According to previous research, manic BD patients had significantly higher relative levels of Eschericha coli and Bifidobacterium adolescentis than depressed BD patients, while depressed people had higher levels of stercoris [51]. Additionally, earlier studies in BD subjects revealed that the bacterial diversity varies between the euthymic and depressive phases, with this finding having an epigenetic effect on the circadian clock gene ARNTL [52]. Regarding ARNTL methylation, illness duration, and gut microbiome diversity, there is an unfavorable correlation [52]. The microbial bacteria Faecalibacterium abundance was negatively correlated with self-reported symptoms and depressive severity [53]. Furthermore, among BD patients, smoking and female sex were linked to the presence of Flavonifractor [54]. (Figure 2) Random forest classification models for the diagnosis and prognosis of bipolar disorder were constructed, which verified that gut microbiota could be used as biological indicators for the diagnosis and prognosis of BD [44,45]. Existing evidence has confirmed that the characteristics of the gut microbiota in BD patients have changed, but how the gut microbiota affects brain function and ultimately leads to the occurrence of BD still needs to be further studied.
Neuroimaging endophenotype in BD patients

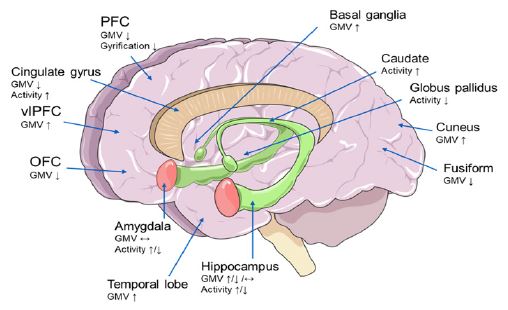
Up until now, MRI technology has made significant advancements in the study of mental illnesses and is frequently used in research on the neuropathological mechanisms and causes of such illnesses. Functional Magnetic Resonance Imaging (fMRI) is an indirect method to evaluate neural activity that uses a real-time blood oxygenation level dependent signal to indicate the degree of regional cerebral blood flow and oxygenation [55]. The Affective Network (AN), Default-Mode Network (DMN), Central-Executive Network (CEN), Salience Network (SN), Frontoparietal Network (FPN), Dorsal Attention Network (DAN), and Ventral Attention Network (VAN) are some of the networks that have received the most attention. The functions of these networks have been discovered to be altered in psychiatric and neurological disorders, including BD [56]. Functional and structural changes in brain regions in BD patients are shown in Figure 3.
Affective-related neural networks in BD patients
Previous studies have demonstrated a close connection between the dysfunction of the emotional processing neural network and the amygdala, Dorsolateral Prefrontal Cortex (dlPFC), Medial Prefrontal Cortex (mdPFC), Ventrolateral Prefrontal Cortex (vlPFC), Anterior Cingulate Cortex (ACC), and Orbitofrontal Cortex (OFC) in mood disorders [57]. During emotional processing and regulation, amygdala activity increased; vlPFC and OFC activity decreased; and the strength of functional connections between the Prefrontal Cortex (PFC) region and the amygdala decreased in the frontal cortex [57]. Functional neuroimaging investigations have revealed abnormalities in the prefrontal cortical-amygdala-centered circuitry for emotion regulation and the prefrontal cortical-striatal reward circuitry in individuals with BD.In BD patients, both the strength of the functional connection between the amygdala and PFC as well as the activity of the emotional network in the vlPFC are greatly reduced. The vlPFC is involved in integrating emotional information, controlling emotional reaction intensity, and controlling autonomic nervous system responses to emotional stimuli. Dysfunction of the vlPFC may make it difficult to control any possible impact areas. For instance, the amygdala is connected to BD mood shift characteristics [57,58]. The amygdala and subcortical system showed an aberrant rise in activity targeted at emotional processing, which may be connected to the emotional instability seen frequently in BD patients [59]. In task-state magnetic resonance investigations, it was discovered that positive stimuli increased the activity of the vlPFC region, whereas negative stimuli lowered the strength of the functional connection between the amygdala and the vlPFC [60]. Previous studies have confirmed that during emotional processing and regulation, there is increased activity of the amygdala and/or decreased activity of the PFC, as well as abnormal functional connectivity between the amygdala and the PFC, and the value of these abnormal neuroimages is positively correlated with the severity of emotional instability, anxiety, depression, and/or mania [61].
Neurocognitive networks in BD patients
The CEN, the DMN and the SN turned out to be the three core neurocognitive networks [56]. According to a meta-analysis of studies using whole-brain seeds to measure resting-state functional connectivity in BD patients, those with acute epiepisodes had lower DMN seed-to-regional connectivity in the PCC and mdPFC than HCs [62]. The DMN seeds and regions of the right dorsal-anterior precuneus in the DAN, as well as parts of the left dlPFC in the FPN, also demonstrated hyperconnectivity in BD patients experiencing acute episodes [62]. When compared to cognitively normal individuals with BD who were in remission, cognitively impaired patients showed hypoactivity in the left dlPFC, frontal and parietal regions of a cognitive control network, as well as hyperactivity in the DMN [63]. According to earlier research, a hyperactive attentional style that prioritizes external stimuli over internal reflection may be the root of the hypoconnectivity between the anterior and posterior DMN in manic BD patients compared to HCs [64]. In contrast, rumination and working memory issues may be linked to the hypoconnectivity in the posterior DMN in depressive BD patients compared to HCs [65]. In BD patients, a different study demonstrated that functional connectivity between the basal ganglia and the left CEN dramatically improved, whereas it was significantly reduced between the basal ganglia and the ventral DMN/precuneus [66]. A thorough investigation revealed decreased functional connectivity between the DMN, CEN, and SN at rest [67].
Structural brain alterations in BD patients
According to research on brain structure, BD patients have smaller amygdalas and hippocampuses, as well as less gray and white matter in their PFCs, anterior temporal lobes, and insular cortex [57]. The abnormal Gray Matter Volume (GMV) of the PFC and limbic related brain regions is related to emotional disturbance in BD depressed patients. A crucial part of the brain involved in processing cognitive and emotional information is the PFC. Affective and cognitive impairments are closely related to PFC thinning [68]. Current findings suggest neuroprogressive losses of GMV in PFC among BD patients, which are larger after first onset and with greater severity of the illness and may be associated with particular genetic characteristics [69]. The ACC is involved in cognitive and emotional tasks [70], and reduced GMV of the ACC was found both in longitudinal and cross-sectional studies in BD patients [69]. For other brain structures, increases in GMV of the temporal lobe, basal ganglia and cuneus in BD patients [69].
Functional and structural brain alterations in brain–gut interactions
Short-chain fatty acids, inflammatory factors, neurotransmitters and other metabolites synthesized by the gut microbiota can enter the central system directly or indirectly and activate brain regions related to emotion, thus causing emotional abnormalities [71]. Animal studies have shown that feeding depressed patients’ excrement to germ-free mice results in depressive-like behavior [72]. Furthermore, the use of antibiotics and probiotics significantly changed depression-like behavior in mice [73]. Additional research has demonstrated that mice kept germ-free have less anxiety-like behavior [71,74,75], and that anxiety-like behavior improved once the mice’s natural intestinal microbiota structure was restored [71]. According to a study using magnetic resonance imaging and behavioral tests on germ-free mice, there are several significant differences between healthy and germ-free mice in terms of the morphology and behavior of their brain tissue. This finding suggests that gut flora is crucial for the normal growth and maturation of gray matter in various brain regions, which has significant implications for animal behavior [76]. Additionally, data from human studies demonstrates that the ACC, through its various subre gions, regulates emotion, behavior, the stomach and other visceral reactions. To regulate emotions, the insula combines emotional information with information from the viscera and the somatosensory system. It is connected to the control of higher order emotions and travels from the insula to the amygdala, hypothalamus, periaquaductal gray matter, and other brain stem locations. PFCS, including OFCs and dlPFC, play an important role in the regulation of emotional and visceral reactions [77]. Furthermore, two investigations split healthy groups into two groups at random. The subjects had fMRI scans before and after receiving probiotic treatment in the experimental group. Both investigations indicated that the activity of brain regions involved in central processing that regulate emotions and feelings could be seen in the probiotic group’s MRI scans, indicating the role of gut flora in the control of brain function [78,79]. Studies have shown that changes in gut flora can affect changes in brain structure, such as changes in gray matter density and volume, cortical thickness, surface area, etc., in addition to their effects on brain function [80]. Taken together, these findings offer significant direct evidence for emotional control of gut sensory inputs at the level of brain anatomy and function. To characterize the microbial-gut-brain axis in BD, recent studies using gut microbial metagenomics, serum metabolomics, and neuroimaging revealed the discovery of BD-related neuroactive microbes and metabolites as potential markers associated with BD-typical features of functional connectivity of brain networks [47]. This study found that some connection networks, such as the functional connectivity of the language areas, thalamus and striatum, sensorimotor areas, hippocampus and amygdala, were related to microorganisms that were associated with neuroactive metabolites in the blood [47].
Limitations of previous studies
Current studies have demonstrated that characteristic changes in gut microbiota, and abnormal structure and function of neural networks, and there is a close correlation between gut microbiota and abnormal structure and function of neural networks. The cause-and-effect relationship between the BD and brain–gut interactions is still not obvious, though. There are few studies that categorize the disease status of BD patients, regardless of gut microbiota or brain function studies in the condition. Patients may have variable gut microbial compositions and functional brain imaging depending on their mood condition. More studies with larger participant samples are required because there are hardly any studies that provide repeated results. Future research should go further to elucidate the relationships between gut microbiota and brain function in BD patients using multiomics approaches such as proteomics, metabonomics, radiomics, and metagenomics.
Conclusion
There were significant alterations in the gut microbiota in BD patients, including decreased microbial diversity and composition changes, which may be related to disease phase and type. Additionally, affective and cognitive-related neural networks involving the AN, DMN, CEN, SN and other networks were altered in BD patients. BD-related microorganisms and metabolites were discovered to be connected to BD-typical brain network functional connectivity properties. This review presents investigations of the link between gut microbiota and neuroimaging in BD patients to further explore the microbiota-gut-brain axis mechanism of BD.
References
- Yatham L, Kennedy S, Parikh S, et al. Canadian Network for Mood and Anxiety Treatments (CANMAT) and International Society for Bipolar Disorders (ISBD) 2018 guidelines for the management of patients with bipolar disorder. Bipolar disorders. 2018; 20: 97-170.
- Vieta E, Berk M, Schulze T, et al. Bipolar disorders. Nature reviews Disease primers. 2018; 4: 18008.
- Pereira A, Oliveira J, Silva S, et al. Inflammation in Bipolar Disorder (BD): Identification of new therapeutic targets. Pharmacological research. 2021; 163: 105325.
- Naaldijk Y, Bittencourt M, Sack U, et al. Kinins and microglial responses in bipolar disorder: a neuroinflammation hypothesis. Biological chemistry. 2016; 397: 283-296.
- Fung TC, Olson CA, Hsiao EY. Interactions between the microbiota, immune and nervous systems in health and disease. Nat Neurosci. 2017; 20: 145-155.
- Gill SR, Pop M Fau - Deboy RT, Deboy Rt Fau - Eckburg PB, et al. Metagenomic analysis of the human distal gut microbiome.
- Leviatan S, Segal E. Identifying gut microbes that affect human health. Nature. 2020; 587: 373-374.
- Keunen K, van Elburg RM, van Bel F, et al. Impact of nutrition on brain development and its neuroprotective implications following preterm birth. Pediatric research .2015; 77: 148-155.
- Cryan JF, O’Riordan KJ, Cowan CSM, et al. The Microbiota-Gut-Brain Axis. Physiol Rev. 2019; 99: 1877-2013.
- Rhee SH, Pothoulakis C, Mayer EA. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat Rev Gastroenterol Hepatol. 2009; 6: 306-314.
- Osadchiy V, Martin CR, Mayer EA. The Gut-Brain Axis and the Microbiome: Mechanisms and Clinical Implications. Clinical gastroenterology and hepatology : the official clinical practice journal of the American Gastroenterological Association. 2019; 17: 322-332.
- Straub RH, Wiest R, Strauch UG, et al. The role of the sympathetic nervous system in intestinal inflammation. Gut. 2006; 55: 1640-1649.
- Lyte M. Microbial endocrinology in the microbiome-gut-brain axis: how bacterial production and utilization of neurochemicals influence behavior. PLoS Pathog. 2013; 9: e1003726.
- Mittal R, Debs LH, Patel AP, et al. Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J Cell Physiol. 2017; 232: 2359-2372.
- Carlson A, Xia K, Azcarate-Peril M, et al. Infant Gut Microbiome Associated With Cognitive Development. Biological psychiatry. 2018; 83: 148-159.
- Cong X, Xu W, Romisher R, et al. Gut Microbiome and Infant Health: Brain-Gut-Microbiota Axis and Host Genetic Factors. The Yale journal of biology and medicine. 2016; 89: 299-308.
- Wang S, Harvey L, Martin R, et al. Targeting the gut microbiota to influence brain development and function in early life. Neuroscience and biobehavioral reviews. 2018; 95: 191-201.
- Babenko O, Kovalchuk I, Metz G. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neuroscience and biobehavioral reviews. 2015; 48: 70-91.
- O’Mahony SM, Marchesi JR, Scully P, et al. Early life stress alters behavior, immunity, and microbiota in rats: implications for irritable bowel syndrome and psychiatric illnesses. Biol Psychiatry. 2009; 65: 263-267.
- Bailey M, Coe C. Maternal separation disrupts the integrity of the intestinal microflora in infant rhesus monkeys. Developmental psychobiology. 1999; 35: 146-155.
- Bailey M, Dowd S, Galley J, et al. Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain, behavior, and immunity. 2011; 25: 397-407.
- Galley J, Bailey M. Impact of stressor exposure on the interplay between commensal microbiota and host inflammation. Gut microbes. 2014; 5: 390-396.
- Dalile B, Van Oudenhove L, Vervliet B, et al. The role of short-chain fatty acids in microbiota-gut-brain communication. Nature reviews Gastroenterology & hepatology. 2019; 16: 461-478.
- Li H, Sun J, Wang F, et al. Sodium butyrate exerts neuroprotective effects by restoring the blood-brain barrier in traumatic brain injury mice. Brain research. 2016; 1642: 70-78.
- Parker A, Fonseca S, Carding S. Gut microbes and metabolites as modulators of blood-brain barrier integrity and brain health. Gut microbes. 2020; 11: 135-157.
- Braniste V, Al-Asmakh M, Kowal C, et al. The gut microbiota influences blood-brain barrier permeability in mice. Science translational medicine. 2014; 6: 263 ra 158.
- Jakobsson H, Rodríguez-Piñeiro A, Schütte A, et al. The composition of the gut microbiota shapes the colon mucus barrier. EMBO reports 2015; 16: 164-177.
- Lutgendorff F, Nijmeijer R, Sandström P, et al. Probiotics prevent intestinal barrier dysfunction in acute pancreatitis in rats via induction of ileal mucosal glutathione biosynthesis. PloS one 2009; 4: e4512.
- Obrenovich M. Leaky Gut, Leaky Brain? Microorganisms. 2018; 6.
- Huang Z, Wong L, Su Y, et al. Blood-brain barrier integrity in the pathogenesis of Alzheimer’s disease. Frontiers in neuroendocrinology 2020; 59: 100857.
- Misiak B, Łoniewski I, Marlicz W, et al. The HPA axis dysregulation in severe mental illness: Can we shift the blame to gut microbiota? Progress in neuro-psychopharmacology & biological psychiatry 2020; 102: 109951.
- Frankiensztajn L, Elliott E, Koren O. The microbiota and the hypothalamus-pituitary-adrenocortical (HPA) axis, implications for anxiety and stress disorders. Current opinion in neurobiology 2020; 62: 76-82.
- Crumeyrolle-Arias M, Jaglin M, Bruneau A, et al. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology 2014; 42: 207-217.
- Desbonnet L, Garrett L, Clarke G, et al. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience. 2010; 170: 1179-1188.
- Gareau M, Jury J, MacQueen G, et al. Probiotic treatment of rat pups normalises corticosterone release and ameliorates colonic dysfunction induced by maternal separation. Gut 2007; 56: 1522-8.
- de Weerth C. Do bacteria shape our development? Crosstalk between intestinal microbiota and HPA axis. Neuroscience and biobehavioral reviews. 2017; 83: 458-471.
- Mawe G, Hoffman J. Serotonin signalling in the gut--functions, dysfunctions and therapeutic targets. Nature reviews Gastroenterology & hepatology. 2013; 10: 473-486.
- De Vadder F, Grasset E, Mannerås Holm L, et al. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proceedings of the National Academy of Sciences of the United States of America. 2018; 115: 6458-6463.
- Clarke G, Grenham S, Scully P, et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Molecular psychiatry. 2013; 18: 666-673.
- Xu R, Wu B, Liang J, et al. Altered gut microbiota and mucosal immunity in patients with schizophrenia. Brain Behav Immun 2019.
- Jiang H, Ling Z, Zhang Y, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain, behavior, and immunity. 2015; 48: 186-194.
- Painold A, Mörkl S, Kashofer K, et al. A step ahead: Exploring the gut microbiota in inpatients with bipolar disorder during a depressive episode. Bipolar disorders. 2019; 21: 40-49.
- Rong H, Xie XH, Zhao J, et al. Similarly in depression, nuances of gut microbiota: Evidences from a shotgun metagenomics sequencing study on major depressive disorder versus bipolar disorder with current major depressive episode patients. J Psychiatr Res. 2019; 113: 90-99.
- Hu S, Li A, Huang T, et al. Gut Microbiota Changes in Patients with Bipolar Depression. Advanced science (Weinheim, Baden-Wurttemberg, Germany). 2019; 6: 1900752.
- Zheng P, Yang J, Li Y, et al. Gut Microbial Signatures Can Discriminate Unipolar from Bipolar Depression. Advanced science (Weinheim, Baden-Wurttemberg, Germany). 2020; 7: 1902862.
- McIntyre R, Subramaniapillai M, Shekotikhina M, et al. Characterizing the gut microbiota in adults with bipolar disorder: a pilot study. Nutritional neuroscience. 2021; 24: 173-180.
- Li Z, Lai J, Zhang P, et al. Multi-omics analyses of serum metabolome, gut microbiome and brain function reveal dysregulated microbiota-gut-brain axis in bipolar depression. Mol Psychiatry. 2022; 27: 4123-4135.
- Huang T, Lai J, Du Y, et al. Current Understanding of Gut Microbiota in Mood Disorders: An Update of Human Studies. Frontiers in genetics. 2019; 10: 98.
- Lai WT, Zhao J, Xu SX, et al. Shotgun metagenomics reveals both taxonomic and tryptophan pathway differences of gut microbiota in bipolar disorder with current major depressive episode patients. J Affect Disord. 2021; 278: 311-319.
- Jiang HY, Xu LL, Zhang X, et al. The microbiome in bipolar depression: A longitudinal study of one pair of monozygotic twins. Bipolar Disord. 2019; 21: 93-97.
- Guo L, Ji, C, Ma, Q, et al. The diversity and the abundance of gut microbiome in patients with bipolar disorder. Chinese Journal of Psychiatry. 2018; 51: 98-104.
- Bengesser SA, Morkl S, Painold A, et al. Epigenetics of the molecular clock and bacterial diversity in bipolar disorder. Psychoneuroendocrinology. 2019; 101: 160-166.
- Evans SJ, Bassis CM, Hein R, et al. The gut microbiome composition associates with bipolar disorder and illness severity. J Psychiatr Res. 2017; 87: 23-29.
- Coello K, Hansen TH, Sorensen N, et al. Gut microbiota composition in patients with newly diagnosed bipolar disorder and their unaffected first-degree relatives. Brain Behav Immun. 2019; 75: 112-118.
- Ogawa S, Lee T, Kay A, et al. Brain magnetic resonance imaging with contrast dependent on blood oxygenation. Proceedings of the National Academy of Sciences of the United States of America. 1990; 87: 9868-9872.
- Menon V. Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn Sci. 2011; 15: 483-506.
- Phillips ML, Swartz HA. A critical appraisal of neuroimaging studies of bipolar disorder: toward a new conceptualization of underlying neural circuitry and a road map for future research. Am J Psychiatry. 2014; 171: 829-843.
- Townsend JD, Torrisi SJ, Lieberman MD, et al. Frontal-amygdala connectivity alterations during emotion downregulation in bipolar I disorder. Biol Psychiatry. 2013; 73: 127-135.
- Phillips ML, Vieta E. Identifying functional neuroimaging biomarkers of bipolar disorder: toward DSM-V. Schizophr Bull. 2007; 33: 893-904.
- Singh MK, Chang KD, Kelley RG, et al. Early signs of anomalous neural functional connectivity in healthy offspring of parents with bipolar disorder. Bipolar Disord. 2014; 16: 678-689.
- Acuff HE, Versace A, Bertocci MA, et al. Association of Neuroimaging Measures of Emotion Processing and Regulation Neural Circuitries With Symptoms of Bipolar Disorder in Offspring at Risk for Bipolar Disorder. JAMA Psychiatry 2018; 75: 1241-1251.
- Wang Y, Gao Y, Tang S, et al. Large-scale network dysfunction in the acute state compared to the remitted state of bipolar disorder: A meta-analysis of resting-state functional connectivity. EBioMedicine. 2020; 54: 102742.
- Zarp Petersen J, Varo C, Skovsen C, et al. Neuronal underpinnings of cognitive impairment in bipolar disorder: A large data-driven functional magnetic resonance imaging study. Bipolar disorders. 2022; 24: 69-81.
- Magioncalda P, Martino M, Conio B, et al. Functional connectivity and neuronal variability of resting state activity in bipolar disorder--reduction and decoupling in anterior cortical midline structures. Human brain mapping. 2015; 36: 666-682.
- Gong J, Chen G, Jia Y, et al. Disrupted functional connectivity within the default mode network and salience network in unmedicated bipolar II disorder. Progress in neuro-psychopharmacology & biological psychiatry. 2019; 88: 11-18.
- Teng S, Lu C, Wang P, et al. Classification of bipolar disorder using basal-ganglia-related functional connectivity in the resting state. Annual International Conference of the IEEE Engineering in Medicine and Biology Society IEEE Engineering in Medicine and Biology Society Annual International Conference 2013; 2013: 1057-1060.
- Yoon S, Kim T, Kim J, et al. Altered functional activity in bipolar disorder: A comprehensive review from a large-scale network perspective. Brain and behavior 2021; 11: e01953.
- Caligiuri MP, Brown GG, Meloy MJ, et al. An fMRI study of affective state and medication on cortical and subcortical brain regions during motor performance in bipolar disorder. Psychiatry Res. 2003; 123: 171-182.
- Lim C, Baldessarini R, Vieta E, et al. Longitudinal neuroimaging and neuropsychological changes in bipolar disorder patients: review of the evidence. Neuroscience and biobehavioral reviews. 2013; 37: 418-435.
- Bush G, Luu P, Posner M. Cognitive and emotional influences in anterior cingulate cortex. Trends in cognitive sciences. 2000; 4: 215-222.
- Clarke G, Grenham S, Scully P, et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry . 2013; 18: 666-673.
- Zheng P, Zeng B, Zhou C, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry 2016; 21: 786-796.
- Yang Z, Li J, Gui X, et al. Updated review of research on the gut microbiota and their relation to depression in animals and human beings. Mol Psychiatry. 2020; 25: 2759-2772.
- Neufeld KM, Kang N, Bienenstock J, et al. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol Motil. 2011; 23: 255-264, e119.
- Arentsen T, Raith H, Qian Y, et al. Host microbiota modulates development of social preference in mice. Microb Ecol Health Dis. 2015; 26: 29719.
- Lu J, Synowiec S, Lu L, et al. Microbiota influence the development of the brain and behaviors in C57BL/6J mice. PLoS One.. 2018; 13: e0201829.
- Van Oudenhove L, Coen SJ, Aziz Q. Functional brain imaging of gastrointestinal sensation in health and disease. World J Gastroenterol. 2007; 13: 3438-3445.
- Tillisch K, Labus J, Kilpatrick L, et al. Consumption of fermented milk product with probiotic modulates brain activity. Gastroenterology. 2013; 144: 1394-401, 401.e1-4.
- Bagga D, Reichert JL, Koschutnig K, et al. Probiotics drive gut microbiome triggering emotional brain signatures. Gut Microbes. 2018; 9: 486-496.
- Tillisch K, Labus J. Neuroimaging the microbiome-gut-brain axis. Advances in experimental medicine and biology. 2014; 817: 405-416.