
Journal of Clinical Images and Medical Case Reports
ISSN 2766-7820
Review Article - Open Access, Volume 4
Small but powerful: The role of HSP in bacterial infectious disease pathogenesis- A mini-review of recent findings
Anekpo Chijioke Chinedu1*, Okpara Titus Chukwubuzo2, Zikora Kizito Glory Anyaegbunam3,4
1Department of Ear, Nose & Throat (ENT), College of Medicine, Enugu State University of Science and Technology, Enugu, Nigeria.
2Department of Internal Medicine, College of Medicine, Enugu State University of Science & Technology, Enugu, Nigeria.
3Department of Microbiology, Faculty of Biological Sciences, University of Nigeria Nsukka, Nigeria.
4Institute for Drug-Herbal Medicine-Excipient-Research and Development, Faculty of Pharmaceutical Sciences, University of Nigeria, Nsukka, Nigeria.
*Corresponding Author : Anekpo Chijioke Chinedu
Department of Ear, Nose & Throat (ENT), College of Medicine, Enugu State University of Science and Technology, Enugu, Nigeria.
Email: Chijioke.anekpo@esut.edu.ng
Received : Sep 26, 2023
Accepted : Oct 20, 2023
Published : Oct 27, 2023
Archived : www.jcimcr.org
Copyright : © Chinedu AC (2023).
Abstract
Heat Shock Proteins (Hsp) are evolutionarily conserved ubiquitous molecular chaperones usually produced when cells are exposed to temperatures beyond their normal growth temperature. They are a group of ATP-independent chaperons pivotal for cell survival and proliferation. This group of proteins has been well studied over the years. However, recent investigations leveraging the available Whole Genome Sequencing (WGS) approach are beginning to shed more light on how this protein interacts with other intracellular proteins to modulate bacterial virulence and pathogenesis. Bacteria cells that manufacture HSPs can survive at higher temperatures. Certain heat-shock proteins have a refolding or degradation action that can save the bacterial cell even in extreme stress demands and even help some bacteria transition from being commensal to a pathogen. Certain HSPs are virulence factors, while others modulate bacterial pathogenesis increasing bacterial resistance to host immunological attack. Interestingly, others also influence certain virulence gene expressions. Several mechanisms govern heat shock response. Moreover, the employment of specific regulatory proteins that exert a positive or negative effect on the initiation of transcription by the RNA polymerase enzyme is successful and efficient in controlling heat-shock gene transcription. Interestingly, most pathogens use several approaches to reprogram gene transcription in response to diverse stress rapidly. In this mini-review, we tactfully discussed the recent update on HSP in bacteria virulence and how these proteins could serve as potential drug targets. Moreover, we provided an overview of how HSPs are regulated in bacterial pathogens favouring bacterial survival in diverse cellular niches.
Keywords: Heat shock proteins (Hsp); Bacteria; Virulence; Pathogenesis.
Citation: Chinedu AC, Chukwubuzo OT, Anyaegbunam ZKG. Small but powerful: The role of HSP in bacterial infectious disease pathogenesis- A mini-review of recent findings. J Clin Images Med Case Rep. 2023; 4(10): 2661.
Introduction
Ritossa originally discovered the heat-shock reaction in the polytene chromosomes of salivary glands of drosophila melanogaster flew when they were shifted from their normal development temperature to 37°C [1]. Ever since that discovery, a lot has been done to understand this group of proteins. Heat Shock Proteins (HSP) have been found in various organisms and play an important function in cell homeostasis. They are a class of proteins induced by high temperatures, meaning they are only synthesized when needed and are influenced by rising temperatures. The key members of this group are a class of functionally related proteins that help other proteins fold and refold. When cells are exposed to high temperatures, HSP expression increases. HSP70 and HSP90 are two chaperones essential for folding and regulating a range of proteins [2].
HSPs are involved in many aspects of protein interactions, including folding, establishing an appropriate protein compound, and preventing undesired protein aggregation [3]. The heat-shock response is an important protective mechanism for bacterial survival and adaptability to harsh environments. This response appears ubiquitous, as it has been detected in all the bacteria studied [3]. It’s made up of a series of well-coordinated responses and processes, the majority of which revolve around the tightly controlled creation of several heat-shock proteins. These, in turn, mostly consist of molecular chaperones and proteases, whose intracellular abundance increases fast in response to a range of environmental challenges. Heat-shock proteins are primarily responsible for assisting protein folding. They also help denatured proteins and harmful misfolded aggregates to be rescued or degraded [4,5].
Furthermore, a change in a bacteria’s environment necessitates a change in behaviour. Bacteria, in particular, have evolved regulatory techniques that combine transcriptional and posttranscriptional processes to rapidly activate heat-shock protein synthesis just when it is needed. To respond quickly to environmental stimuli and guarantee that heat-shock proteins are only expressed, when necessary, bacteria use sensory biomolecules that can detect temperature variations and translate them into coordinated gene expression outputs [6]. Different microorganisms have developed a diverse set of heat sensors in this regard.
Some heat-shock proteins are key virulence factors, while others appear to affect pathogenesis indirectly, in addition to their responsibilities in protecting cellular proteins from environmental insults and maintaining cellular homeostasis. Even though the heat-shock response is conserved across all prokaryotes and eukaryotes, the basic molecular processes governing heat-shock regulation are not completely understood in different bacterial species considering that genes differ considerably among bacterial species [5]. In this mini-review, we discussed the recent update on the involvement of HSP in virulence and pathogenesis mechanisms in bacterial pathogens. Furthermore, we provided an overview of recent findings on the molecular interplay and cross-talk between HSP and virulence genes. This is very critical to properly understand the role of HSP virulence and how it could be exploited to develop better therapeutic development.
An overview of HSPs in bacteria
Physiological stresses (such as heat, hemodynamics, mutant proteins, and oxidative damage) cause various changes in a cell, affecting protein structures and function. As a defence strategy to maintain survival, bacterial pathogens launch a cascade of processes that engage critical proteins, the molecular chaperones, in an approach to repair or degrade damaged proteins. Molecular chaperones like the Heat Shock family of stress Proteins (HSPs) have been shown to have a role in a variety of cellular activities. Stress-inducible regulatory networks, both at the transcriptional and posttranscriptional levels, add to the adaptability of the ubiquitous HSP family. Two key lines of data provide insight into the mechanisms underpinning HSP function: (1) Protein-folding machinery, or molecular chaperones, are required for the correct folding of many proteins in a cell, and (2) HSP chaperones repair denatured proteins or promote their destruction following heat shock.
Generally, the heat-shock response, which is triggered by environmental stress shocks, involves the induction or upregulation of the production of a collection of proteins that enable the cell to survive in conditions that would ordinarily kill it [5]. The heat-shock response is a cellular defence mechanism that rapidly synthesises numerous heat-shock proteins in response to sudden adverse environmental growth conditions. Temperature changes are sensed by particular sensory biomolecules in bacteria, transmitting intercellular signals that coordinate gene expression outputs. Nucleic acids (DNA or RNA) and proteins are examples of sensory biomolecules, often known as thermosensors [4]. When a stress signal is detected, it is translated into specific molecular pathways that control the transcription of heat-shock protein genes. Heat-shock gene transcription can be regulated in either a positive or negative manner by specialised regulatory proteins. Positive regulation uses alternate sigma factors to drive the RNA polymerase enzyme to a subset of selected promoters, whereas transcriptional repressors are responsible for negative control. Interestingly, whereas some bacteria use just positive or negative processes, these two opposing techniques coexist in some microorganisms, forming complex networks that regulate heat-shock genes.
When bacteria cells are exposed to higher temperatures, heat-shock proteins (hsps) are induced quickly and transiently to deal with increased protein damage. The mechanism underpinning hsps induction has been a major research focus in several quarters. The cellular level of sigma 32, which is important for heat-shock gene transcription, increases fast and transiently when the temperature rises [7]. A recent study of translational induction and subsequent shut-off of sigma 32 synthesis provided a clue to a better understanding of early regulatory events.
Important HSPs in bacteria
HSP70
Hsp70 proteins anchor the cellular network of molecular chaperones and folding catalysts. By transiently associating their substrate binding domain with short hydrophobic peptide segments inside their substrate proteins, they aid a wide range of protein folding activities in the cell. Switching Hsp70 between the low-affinity ATP bound state and the high-affinity ADP bound state drives the substrate binding and release cycle. In vitro and in vivo, ATP binding and hydrolysis are required for Hsp70 proteins to function as chaperones. Co-chaperones of the J-domain protein family, which target Hsp70s to their substrates, and nucleotide exchange factors, which determine the lifetime of the Hsp70-substrate complex, control the ATPase cycle [8]. Hsp70s help with a variety of folding activities, including the folding and assembly of newly produced proteins, the refolding of misfolded and aggregated proteins, membrane transfer of organellar and secretory proteins, and regulatory protein activity control. Hsp70s have both housekeeping and quality control functions in the cell, as they are built-in components of folding and signal transduction pathways [8]. They proofread protein structure and repaired misfolded conformers. These groups of proteins have the ability to bind with hydrophobic peptide regions of proteins in an ATP-controlled manner.
HSP90
HSP90 has kinase activity and can bind to nuclear receptor ligands through an unknown route. HSP90 is a molecular chaperone that promotes protein folding, refolding, and activation across a wide range of substrates. HSP90 can interact with unfolded substrates that are late in their folding route [9]. HSP90 has the ability to actively affect the binding of substrates, altering the substrate folding process and resulting in a functional. Bacterial HSP90 is critical in protein folding in aberrant situations such as heat stress. The N-terminal domains of HSP90 need ATP to deform and then make contact with the middle domain. HSP90 only returns to the open state if ATP is used. HSP90 is a macromolecular protein assembly group member, which plays a key role in scaffolding proteins. The N-terminal, middle, and C-terminal domains make up the HSP90 dimer [8].
Chaperones may fold proteins, which is their primary job in ensuring proper assembly. When the temperature is lowered, these proteins primarily regulate protein production. DnaK and GroEL are the two major chaperones linked to HSP90 [10,11]. HSP90 interacts with the DnaK (HSP70) chaperone. GroEL (HSP60) chaperone is required for development in a variety of environments [11]. DnaK and GroEL: Major HSPs (DnaK, GroEL) are molecular chaperones involved in DNA replication, UV mutagenesis, bacterial growth, RNA transcription, and flagella formation, among other cellular functions. GroEL and GroES work cooperatively to translocate proteins over membrane barriers, allowing secretion. These HSPs are responsible for preventing protein denaturation and reactivating partially denatured proteins. On the other hand, DnaK is a chaperone that utilises ATP and ADP released by the N-terminal domain, as well as ATP catalysis and the release of unfolded proteins by the C-terminal domain. DnaK can bind to nascent polypeptide chains and aid in the refolding and destruction of proteins [6].
An overview of heat sensing mechanisms
Heat-sensing mechanisms that integrate environmental cues to activate suitable response pathways are required for bacteria to respond quickly to unexpected temperature increases. Multiple thermoregulation processes in bacteria have been reported, using practically all kinds of biomolecules, including lipids, proteins, and nucleic acids (i.e., DNA and RNA). All of these classes can function as thermosensors, detecting changes in the ambient temperature and triggering appropriate cellular responses (Figure 1). Therefore, heat-sensing systems can be either direct, in which the temperature affects the activity of the detecting biomolecule directly, or indirect, in which the repercussions of a fast temperature increase (for example, the aggregation of mismisfolded proteins in the cytoplasm) are observed. Even though the temperature is a ubiquitous signal that regulates various cellular pathways, several thermosensors control virulence determinants [12,13].
Temperature can be sensed through RNA. Because temperature influences the translation efficiency of both the intracellular pool of mRNA molecules and already-in-progress transcription, this thermoregulation mechanism ensures a very quick response upon signal reception. The main premise is based on the creation of zipper-like, temperature-sensitive secondary structures in mRNAs that are regulated in this way [14-16]. Temperature can also be sensed through the DNA of the bacterial cell in some situations [17]. The environmental factors experienced in a microorganism’s ecological habitat influence several physiological and metabolic pathways. As a result, the cell’s metabolic status, particularly the ADP/ATP ratio, is affected. Enzymes that require ATP as a cofactor, such as DNA gyrase, will be impacted as a result. This enzyme, whose activity is dependent on ATP and inhibited by ADP, is dependent on ATP and inhibited by ADP (so change in the ATP: ADP ratio influences DNA gyrase activity). As a result, external fluctuating variables that alter metabolic processes, such as osmotic and thermal shock, can have an impact on the global degree of DNA supercoiling [18,19]. Because DNA supercoiling can affect gene transcription, it can be thought of as a thermosensor for the environmental temperature change that responds to external stimuli by changing the global topological state of the chromosome [20]. Plasmid supercoiling is one of the most important DNA topological parameters that change with temperature. It has been shown that abrupt temperature fluctuations cause transitory changes in plasmid DNA architecture, which has a major impact on transcription efficiency in certain bacterial pathogens [21].
Finally, bacteria can use proteins as heat sensors to detect and respond to temperature fluctuations in their ecological niches. Temperature sensors have been identified in a variety of proteins, including kinases, heat-shock transcriptional repressors, and chaperones [12]. On the other hand, the two latter kinds of proteins are principally used to translate stress signals into a transcriptional heat-shock response. These thermosensing repressors undergo a conformational shift in response to heat shock, lowering their relative binding affinity for their operators. As a result, target gene transcription is unrestricted. CtsR, the global repressor of heat-shock genes in Bacillus subtilis and other low-GC Gram-positive bacteria, is one of the best-studied examples [22].
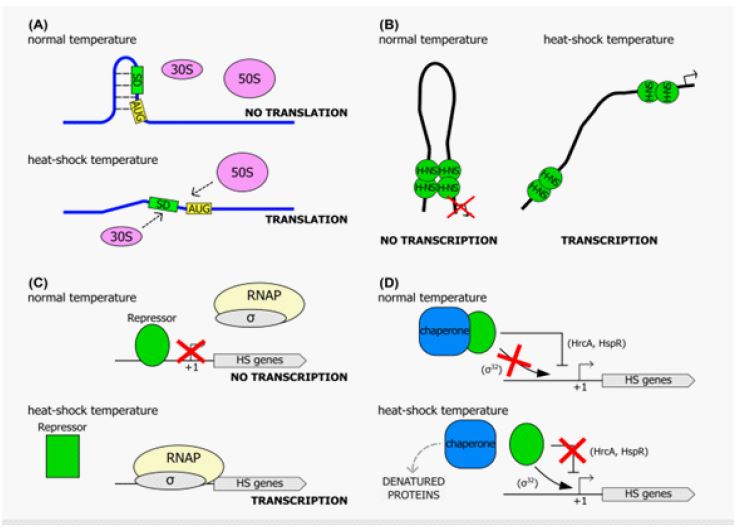
The temperature-dependent conformational change that causes the loss of DNA-binding ability in CtsR has been demonstrated to be restricted to a small glycine-rich loop region within the DBD, which serves as the exact functional site for heat sensing [22]. RheA, a heat-shock repressor from Streptomyces albus, has been reported as capable of sensing temperature fluctuations without other variables and functioning similarly to CtsR. RheA loses its DNA-binding function when exposed to heat, and the transition from active to inactive was linked to a temperature-induced reversible conformational change.
Regulation of heat shock proteins in bacteria
Alternative sigma factors drive RNA polymerase to particular promoters that differ from housekeeping promoters, allowing HSP to be regulated. Sigma-32 and sigma-E (E) are two prominent sigma factors that have been discovered [23]. The rpoH gene encodes the sigma-32 factor, which is involved in protein stability and translation efficiency and is controlled post-transcriptionally. Under typical conditions, the amount of active sigma-32 is kept to a minimum [24].
First, increasing the temperature from 300 to 39°C or higher causes the rpoH gene to translate faster. Second, temperature-sensitive secondary structures arise within the five’ region of the rpoH gene that detach the ribosome-binding site, but these secondary structures deliquesce at higher temperatures, allowing for a more effective translation of the rpoH message. Most HSPs are generated at low levels under normal settings, but when exposed to high temperatures, they are induced quickly and transiently. Finally, heat-shock gene regulation in bacteria can be positive or negative [25]. Transcriptional repressors mediate negative regulation, while positive regulation uses alternate sigma factors to direct the transcriptional machinery to a subset of specified promoters. Interestingly, whereas some bacteria use just positive or negative processes, these opposing techniques coexist in some species, forming a complex regulatory network of heat-shock genes [26].
The employment of a specialised alternative sigma factor, a subunit of the RNA polymerase that imparts promoter recognition specificity to the transcription enzyme, is required for positive transcriptional control of heat-shock genes. This transcriptional regulation mechanism takes advantage of alternative factors’ ability to outcompete the housekeeping subunit ordinarily associated with RNA polymerase, redirecting the enzyme to a subset of heat-shock gene promoters and reprogramming cellular transcription. Using a specialised factor to transiently increase the transcription of heat-shock genes dependent on temperature stress is frequently utilised among bacteria, with Escherichia coli serving as an excellent example. Table 1 summarises the regulatory mechanisms controlling HSPs gene transcription in several bacterial pathogens as [6] described.
The role of HSPs in bacterial pathogenesis
Several stressors, including temperature changes, osmotic shifts, desiccation, antibiotics, solvents, or heavy metals, might trigger a heat-shock response. However, the heat-shock response, which is caused by a rapid increase in temperature, has long been utilised as a model system for investigating the effects of stress on biological systems. Environmental stress has a major impact on cellular protein homeostasis. Heat shock proteins aid in folding newly generated proteins, inhibit protein aggregation under stress circumstances and recover proteins that have been partially or totally unfolded by stresses such as a fast temperature increase in bacterial cells. In fact, during times of stress, the spontaneous folding of newly synthesised proteins is inefficient and error-prone, and chaperones fold the majority of nascent polypeptides. In addition, a high proportion of already folded proteins denature partially or entirely, making them more prone to forming harmful aggregates [27].
Table 1: Regulatory mechanisms controlling HSPs genes transcription in several bacterial pathogens.
| Organisms+ | Transcriptional regulator | Regulated genes |
|---|---|---|
| Bacillussubtilis | HrcA | Class I: groESL, dnaK-dnaJ, grpE, and others |
| σB | Class II: genes coding for generalstress proteins | |
| CtsR | Class III: clpP, clpE, ctsR-mcsA-mcsB-clpC and others | |
| Unknown CssS/CssR | Class IV: htpG | |
| Agrobacteriumtumefaciens | RpoH | groESL,dnaK-dnaJ, grpE, clpP and others |
| HrcA | groESL | |
| Escherichiacoli | σ32 (σH,RpoH) | groESL,dnaK-dnaJ, grpE, ibpA and others |
| σE (σ24) | degP,clpX, lon and others | |
| Listeriamonocytogenes | HrcA | Class I: groESL, dnaK, andothers |
| σB | Class II: genes coding for generalstress proteins | |
| CtsR | Class III: clpP, clpB, hslU and others | |
| Pseudomonasaeruginosa | σH (RpoH) | groESL,dnaK, and others |
| Vibriocholerae | σ32 (RpoH) | groESL,dnaK-dnaJ, lon, clpB, and others |
| Streptococcuspneumoniae | HrcA | groESL,dnaK-dnaJ, grpE |
| Staphylococcusaureus | HrcA | hrcA-dnaK,groESL |
| CtsR | clpP,clpC, clpB, hrcA-dnaK, groESL | |
| σB | genes coding for general stressproteins | |
| Helicobacterpylori | HspR; HrcA | cbpA-hspR-helicase, groESL, hrcA-grpE-dnaK; groESL, hrcA-grpE-dnaK |
Certain heat-shock proteins have been proposed as key virulence factors, whereas others appear to impact pathogenesis indirectly. Molecular chaperones appear to have taken on a variety of activities other than protein folding in this regard, as evidenced by a number of examples. For example, GroEL1, a GroEL paralog with no heat-shock activities [28], is implicated in mycolic acid synthesis and biofilm formation in Mycobacterium smegmatis. Furthermore, Helicobacter pylori GroES homolog performs a role in the storage and trafficking of Ni2+ ions, a critical virulence factor for this human gastric virus, in addition to its archetypal activity as co-chaperonin [3].
Hsp90 play a crucial role in the virulence and pathogenesis of certain bacterial pathogens as reported in several kinds of literature [29-36]. For example, in extraintestinal E. coli (ExPEC) HSP90 has been reported to be playing a critical role. According to a study by [30], it was shown that the production of certain critical virulence factors (e.g., siderophore, yersiniabactin, and colibactin) was disrupted when HSP90 was absent. Also, in Pseudomonas sp, the biosurfactant arthrofactin was reported to be produced by complex pathways, which indirectly suggests the role of HSP90 [37-41]. Many other studies have reported the involvement of HSP90 in virulence in Pseudomonas aeruginosa, Salmonella enterica Typhimurium, Leptospira interrogans, and francisella tularensis [29,42,31-33,36]. HSPs also play crucial roles in protein degradation [43,42,30,44,45].
HSPs as drug targets
An increase in antimicrobial-resistant bacterial pathogens is a major challenge in public health. This has led scientists at different quarters to start exploiting other alternatives in treating multi-drug-resistant pathogens. Certain HSPs like HSPs90 and HSP70 could be new drug targets. For example, research had shown that in E. coli, when the encoding HscA and HscC were deleted, viability was not affected. Targeting these proteins will obviously not lead to growth inhibition. However, it has been shown that the deletion of dnaK leads to filamentous growth. It also resulted in temperature sensitivity above 35°C. However, filamentous growth can be compensated by rpoH gene suppressor mutation. This rpoH encodes the HSP transcription factor σ32.
It is important to emphasise that most pathogens are dependent on DnaK. Thus, the loss of DnaK could definitely affect the cell. The DnaK is crucial for the persistence and cell regrowth of cells in their dormant state [46,47]. Therefore, DnaK could be a target for novel antimicrobial drugs as drugs that could distinguish DnaK from eukaryotic HSP70 could be developed. This will help to prevent pathogen growth without affecting protein homeostasis in eukaryotic cells. Further insight into DnaK was also provided in a study by [48]. Antimicrobial peptides from insects have also been shown to bind DnaK. Through this binding, there was an inhibition of gram-negative pathogens [49]. Similar investigations have also been carried out elsewhere [50,51]. However, it is still not completely clear if the DnaK is the main target of the peptides, as demonstrated in the study by [52,53]. Small molecule activators and inhibitors of DnaK have also been reported [54,55]. however, it is yet to be completely determined if these small molecules only target DnaK or also affect hyman Hsp70.
Due to the increasing resistance crisis, more effort is to be devoted to engineering bacterial Hsp70 inhibitors. A study by [56] showed that chaperone, Hsp90 was responsible for resistant development to specific antimicrobial agents. Overall, more insight is still needed better to understand the utility of this protein as drug targets.
Conclusion
Heat-Shock Proteins (HSPs) have been implicated in various processes, including chaperone action, protein folding, apoptosis and immunology. They have been exploited to generate preventative and therapeutic vaccines for infectious and noninfectious disorders. Several studies have also shown a link between HSPs and treatment resistance and their potential use as a new biomarker for diagnosing many ailments. Bacterial cells that manufacture HSPs can survive at higher temperatures. When the temperature of a bacterial cell is raised, natural proteins are damaged; however, HSPs that have a refolding or degradation action can save the bacterial cell. This impact aids the bacterial cell’s survival throughout its transition from the environment to the body.
Moreover, the heat-shock response, a highly conserved defence mechanism against environmental stress, is governed by a variety of mechanisms in bacteria. To be able to survive in certain niches, bacteria use both positive and negative regulatory mechanisms, either alone or in combination, to rapidly reprogram gene transcription in response to stress and, thus, maintain the internal environment of the bacteria. We have shown that HSPs play critical roles in bacterial virulence and pathogenicity. However, more remains to be done to properly uncover the role of these HSPs in less studied bacterial lineages and to understand their interactions with cellular proteins, how they influence virulence in major bacterial pathogens, and how they could be exploited as good drug targets.
Declarations
Conflict of interest: The authors report no conflict of interest.
Funding: Authors received no funding for this study.
References
- Ritossa F. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia. 1962; 18: 571-573.
- Danielli A, Amore G, Scarlato V. Built shallow to maintain homeostasis and persistent infection: insight into the transcriptional regulatory network of the gastric human pathogen Helicobacter pylori. Nature. 2017; 405: 590-593.
- de Reuse H, Vinella D, Cavazza C. Common themes and unique proteins for the uptake and trafficking of nickel, a metal essential for the virulence of Helicobacter pylori. Front Cell Infection Microbiology. 2018; 3: 94.
- Fiocco D, Collins M, Muscariello L. The Lactobacillus plantarum ftsH gene is a novel member of the CtsR stress response regulon. Journal Bacteriology. 2017; 191: 1688-1694.
- Fiocco D, Capozzi V, Collins M. Characterisation of the CtsR stress response regulon Lactobacillus plantarum. Journal Bacteriology. 2020; 192: 896-900.
- Roncarati D, Danielli A, Scarlato V. CbpA acts as a modulator of HspR repressor DNA binding activity in Helicobacter pylori. Journal Bacteriology. 2015; 193: 5629-5636.
- Roncarati D, Danielli A, Scarlato V. The HrcA repressor is the thermosensor of the heat-shock regulatory circuit in the human pathogen Helicobacter pylori. Molecular Microbiology. 2014; 92: 910-920.
- Zomer A, Fernandez M, Kearney B. An interactive regulatory network controls stress response in Bifidobacterium breve UCC2003. Journal Bacteriology. 2019; 191: 7039-7049.
- Zomer A, vanSinderen D. Intertwinement of stress response regulons in Bifidobacterium breve UCC2003. Gut Microbes. 2018; 1: 100-102.
- Gvozdenov Z, Kolhe J, Freeman BC. The Nuclear and DNA-Associated Molecular Chaperone Network. Cold Spring Harb Perspect Biol. 2019; 11: a034009.
- Grindle MP, Carter B, Alao JP, Connors K, Tehver R, et al. Structural Communication between the E. coli Chaperones DnaK and Hsp90. Int J Mol Sci. 2021; 22: 2200.
- Klinkert B, Narberhaus F. Microbial thermosensors. Cell Molecular Life Science. 2017; 66: 2661-76.
- Shapiro RS, Cowen LE. Thermal control of microbial development and virulence. molecular mechanisms of microbial temperature sensing. Molecular biology. 2018; 3: 238-212.
- Kortmann J, Sczodrok S, Rinnenthal J. Translation on demand by a simple RNA-based thermosensor. Nucleic Acids Resistance. 2016; 39: 2855-2868.
- Roncarati D, Spohn G, Tango N. Expression, purification and characterisation of the membrane-associated HrcA repressor protein of Helicobacter pylori. Protein Expression Purification. 2018; 51: 267-275.
- Waldminghaus T, Gaubig L, Klinkert B. The Escherichia coliibpA thermometer is comprised of stable and unstable structural elements. RNA Biology. 2018; 6: 455-463.
- Nickerson CA, Achberger EC. Role of curved DNA in binding of Escherichia coli RNA polymerase to promoters. Journal Bacteriology. 2018; 177: 5756-5761.
- Hsieh LS, Burger RM, Drlica K. Bacterial DNA supercoiling and [ATP/ADP]. Changes associated with a transition to anaerobic growth. Journal Molecular Biology. 2015; 219: 443-450.
- Dorman CJ, Corcoran CP. Bacterial DNA topology and infectious disease. Nucleic Acids Resistance. 2017; 37: 672-678.
- Colonna B, Casalino M, Fradiani PA. H-NS regulation of virulence gene expression in enteroinvasive Escherichia coli harboring the virulence plasmid integrated into the host chromosome. Journal Bacteriology. 2017; 177: 4703-4712.
- Wilson AC, Tan M. Functional analysis of the heat shock regulator HrcA of Chlamydia trachomatis. Journal Bacteriology. 2020; 184: 201-208.
- Elsholz AK, Michalik S, Zuhlke D. CtsR, the Gram-positive master regulator of protein quality control, feels the heat. Journal bacteriology. 2019; 29: 3621-3629.
- Wang Y, deHaseth PL. Sigma 32-dependent promoter activity in vivo: sequence determinants of the groE promoter. J Bacteriol. 2003; 185: 5800-6.
- Nevozhay D, Adams R, Murphy K. Negative autoregulation linearises the dose-response and suppresses the heterogeneity of gene expression. Post-trancritional Academy Science USA. 2020; 106: 5123-5128.
- Roncarati D, Scarlato V. Regulation of heat-shock genes in bacteria: from signal sensing to gene expression output. FEMS Microbiol Rsev. 2017; 41: 549-574.
- Howell HA, Battista JR. HspR is a global negative regulator of heat shock gene expression in Deinococcus radiodurans. Molecular Microbiology. 2018; 55: 1579-1590.
- Matuszewska M, Kuczynska-Wi´ snik D, Laskowska E. The small heat shock protein IbpA of Escherichia coli cooperates with IbpB in stabilisation of thermally aggregated proteins in a disaggregation competent state. Journal Biochemistry. 2020; 280: 12292-12298.
- Ojha A, Anand M, Bhatt A. GroEL1: A dedicated chaperone involved in mycolic acid biosynthesis during biofilm formation in mycobacteria. Cell biology. 2020; 14: 123-127.
- Dang W, Hu Y-h, Sun L. HtpG is involved in the pathogenesis of Edwardsiella tarda. Vet. Microbiol. 2011; 152: 394-400.
- Garcie C, Tronnet S, Garénaux A, McCarthy AJ, Brachmann AO, et al. The Bacterial Stress-Responsive Hsp90 Chaperone (HtpG) Is Required for the Production of the Genotoxin Colibactin and the Siderophore Yersiniabactin in Escherichia coli. J. Infect. Dis. 2016; 214: 916-24
- Grudniak AM, Klecha B, Wolska KI. Effects of null mutation of the heat-shock gene htpG on the production of virulence factors by Pseudomonas aeruginosa. Future Microbiol. 2018; 13: 69-80.
- King AM, Pretre G, Bartpho T, Sermswan RW, Toma C, et al. High-temperature protein G is an essential virulence factor of Leptospira interrogans. Infect. Immun. 2014; 82: 1123-31.
- Lee M-K, Lee Y, Huh J-W, Chen H, Wu W, et al. The Pseudomonas aeruginosa HSP90-like protein HtpG regulates IL-8 expression through NF-κB/p38 MAPK and CYLD signaling triggered by TLR4 and CD91. Microbes and Infection. 2020.
- Verbrugghe E, Van Parys A, Leyman B, Boyen F, Haesebrouck F, et al. HtpG contributes to Salmonella Typhimurium intestinal persistence in pigs. Vet. Res. 2015; 46: 118.
- Vivien E, Megessier S, Pieretti I, Cociancich S, Frutos R, et al. Xanthomonas albilineans HtpG is required for biosynthesis of the antibiotic and phytotoxin albicidin. FEMS microbiology letters. 2005; 251: 81-9.
- Weiss DS, Brotcke A, Henry T, Margolis JJ, Chan K, et al. In vivo negative selection screen identifies genes required for Francisella virulence. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104: 6037-42.
- Washio K, Lim SP, Roongsawang N, Morikawa M. Identification and characterisation of the genes responsible for the production of the cyclic lipopeptide arthrofactin by Pseudomonas sp. MIS38. Bioscience, Biotechnology, and Biochemistry. 2010; 74: 992-9.
- Huang G, Zhang L, Birch RG. A multifunctional polyketide-peptide synthetase essential for albicidin biosynthesis in Xanthomonas albilineans. Microbiology (Reading). 2001; 147: 631-42.
- Nougayrède J-P, Homburg S, Taieb F, Boury M, Brzuszkiewicz E, et al. Escherichia coli induces DNA double-strand breaks in eukaryotic cells. Science. 2006; 313: 848-51.
- Perry RD, Fetherston JD. Yersiniabactin iron uptake: Mechanisms and role in Yersinia pestis pathogenesis. Microbes and Infection. 2011; 13: 808-17.
- Roongsawang N, Hase K, Haruki M, Imanaka T, Morikawa M, et al. Cloning and characterisation of the gene cluster encoding arthrofactin synthetase from Pseudomonas sp. MIS38. Chem Biol. 2003; 10: 869-80.
- Edkins AL. CHIP: A co-chaperone for degradation by the proteasome. Subcell. Biochem. 2015; 78: 219-42.
- Balasubramanian A, Markovski M, Hoskins JR, Doyle SM, Wickner S. Hsp90 of E. coli modulates assembly of FtsZ, the bacterial tubulin homolog. Proceedings of the National Academy of Sciences of the United States of America. 2019; 116: 12285-94.
- Honoré FA, Maillot NJ, Méjean V, Genest O. Interplay between the Hsp90 Chaperone and the HslVU Protease To Regulate the Level of an Essential Protein in Shewanella oneidensis. MBio. 2019; 10.
- Yosef I, Goren MG, Kiro R, Edgar R, Qimron U. High-temperature protein G is essential for activity of the Escherichia coli clustered regularly interspaced short palindromic repeats (CRISPR)/Cas system. Proceedings of the National Academy of Sciences of the United States of America. 2011; 108: 20136-41.
- Wu CC, Naveen V, Chien CH, Chang YW, Hsiao CD. Crystal Structure of DnaK Protein Complexed with Nucleotide Exchange Factor GrpE in DnaK Chaperone System. J. Biol. Chem. 2012; 287: 21461-21470.
- Pu Y, Li Y, Jin X, Tian T, Ma Q, et al. ATP-dependent Dynamic Protein Aggregation Regulates Bacterial Dormancy Depth Critical for Antibiotic Tolerance. Mol. Cel. 2018; 73: 143-156. e4.
- Hassan AQ, Kirby CA, Zhou W, Schuhmann T, Kityk R, et al. The Novolactone Natural Product Disrupts the Allosteric Regulation of Hsp70. Chem. 2015.
- Scocchi M, Tossi A, Gennaro R. Proline-rich Antimicrobial Peptides: Converging to a Non-lytic Mechanism of Action. Cel. Mol. Life Sci. 2011; 68: 2317-2330.
- Otvos L, Rogers IO ME, Consolvo PJ, Condie BA, et al. (2000). Interaction between Heat Shock Proteins and Antimicrobial Peptides†. Biochemistry. 2000; 39: 14150-14159.
- Zahn M, Berthold N, Kieslich B, Knappe D, Hoffmann R, et al. Structural Studies on the Forward and Reverse Binding Modes of Peptides to the Chaperone DnaK. J. Mol. Biol. 2013; 425: 2463-2479.
- Czihal P, Knappe D, Fritsche S, Zahn M, Berthold N, et al. Api88 Is a Novel Antibacterial Designer Peptide to Treat Systemic Infections with Multidrug-Resistant Gram-Negative Pathogens. ACS Chem. Biol. 2012; 7: 1281-1291.
- Krizsan A, Volke D, Weinert S, Sträter N, Knappe D, et al. Insect-Derived Proline-Rich Antimicrobial Peptides Kill Bacteria by Inhibiting Bacterial Protein Translation at the 70 S Ribosome. Angew. Chem. Int. Ed. 2014; 53: 12236-12239.
- Wisén S, Androsavich J, Evans CG, Chang L, Gestwicki JE. Chemical Modulators of Heat Shock Protein 70 (Hsp70) by Sequential, Microwave-Accelerated Reactions on Solid Phase. Bioorg. Med. Chem. Lett. 2008; 18: 60-65.
- Chang YW, Sun YJ, Wang C, Hsiao CD. Crystal Structures of the 70-kDa Heat Shock Proteins in Domain Disjoining Conformation. J. Biol. Chem. 2008b; 283: 15502-15511.
- Cowen LE, Lindquist S. Hsp90 Potentiates the Rapid Evolution of New Traits: Drug Resistance in Diverse Fungi. Science. 2005; 309: 2185-2189.