
Journal of Clinical Images and Medical Case Reports
ISSN 2766-7820
Review Article - Open Access, Volume 5
Role of the lncRNA HOX antisense intergenic RNA myeloid 1 in Cancer: A Review
Yipei Jing; Shasha Wang; Ye Guo; Xuzhen Qin; Yongzhe Li*
Department of Clinical Laboratory, State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Science and Peking Union Medical College, Beijing, China.
*Corresponding Author : Yongzhe Li
Department of Clinical Laboratory, State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Science and Peking Union Medical College, Beijing, China.
Email: yongzhelipumch@126.com
Received : Mar 12, 2024
Accepted : Apr 02, 2024
Published : Apr 09, 2024
Archived : www.jcimcr.org
Copyright : © Li Y (2024).
Abstract
Long noncoding RNAs (lncRNAs) have emerged as a new family of master regulators of cancer. The HOXA transcript antisense RNA myeloid-specific 1 (HOTAIRM1) located in the HOXA gene cluster was first discovered in acute promyelocytic leukemia. Increasing evidence have showed that HOTAIRM1 is involved in diverse cancer. It regulates tumor proliferation, invasion, and migration via various mechanisms and is related to the clinicopathological characteristics of tumor patients. This article reviews its expression, function, and molecular mechanism in various malignant tumors and future challenges for the diagnosis and treatment of cancers through HOTAIRM1.
Keywords: LncRNA; HOTAIRM1; cancer.
Abbreviations: lncRNAs: Long Noncoding RNAs; HOTAIRM1: HOXA Transcript Antisense RNA Myeloid-Specific 1; AML: Acute Myeloid Leukemia; OS: Osteosarcoma; EC: Endometrial Cancer; EMT: Epithelial–Mesenchymal Transition; TC: Thyroid Cancer; NSCLC: Non-Small Cell Lung Cancer; OSCC: ORAL Squamous Cell Carcinoma; PCa: Prostate Cancer; PDAC: Pancreatic Ductal Adenocarcinoma; HNT: Head and Neck Tumor; HCC: Hepatocellular Carcinoma; ADC: Lung Adenocarcinoma; GC: Gastric Cancer; CRC: Colorectal Cancer; PRC2: Polycomb Repressive Complex 2; H3K27me3: Lysine Residue 27 of Histone 3; spMN: Recapitulates Spinal MN.
Citation: Jing Y, Wang S, Guo Y, Qin X, Li Y. Role of the lncRNA HOX antisense intergenic RNA myeloid 1 in Cancer: A Review. J Clin Images Med Case Rep. 2024; 5(4): 2974.
Introduction
Cancer is a complicated pathological event that can be developed by a number of driving forces and effects including infectious diseases, environmental factors, gene dysregulation and epigenetic alterations [1,2]. Advances in genetic research have revealed the pathogenesis of many cancers, which has led to the development of effective treatments [3,6]. For example, leukemia caused by breakpoint cluster region-Abelson mutations can be effectively cured using imatinib. Protein-coding genes account for < 2% of the total genome. The remaining genome consists of noncoding genes that are responsible for the vast majority of tumorigenesis [7,9].
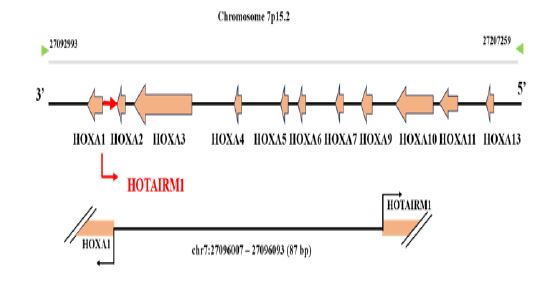
Long noncoding RNAs (lncRNAs) are defined as transcripts longer than 200 nucleotides with little or no predicted protein-coding potential [10,11]. Recent observations have illustrated that a large number of deregulated lncRNAs are involved in human cancers and their function as oncogenes and tumor suppressors in cancer onset and progression [12,15]. HOXA transcript antisense RNA myeloid-specific 1 (HOTAIRM1) is a novel identified lncRNA that maps within the HOXA gene cluster at human chromosome 7p15.2 (Figure 1) [16]. HOTAIRM1, first discovered as an important factor for granulocytic differentiation in NB4 promyelocytic leukemia [17]. Since this discovery, HOTAIRM1 has attracted intense investigation in the field of cancer. A search for HOTAIRM1 and cancer as keywords in the title/abstract yielded 93 articles in PubMed as of December 2023. In this review, we summarized the current knowledge regarding the expression levels, functions, as well as the molecular mechanism underlying HOTAIRM1 regulation on cancer hallmarks.
HOTAIRM1 expression in cancer: Multiple studies have been reported that lncRNAs are aberrantly expressed in various diseases, especially in malignant tumors [18,20]. In general, lncRNAs dysregulation contribute to tumors development by promotion, proliferation, invasion and metastasis of tumor cells [21,23]. LncRNA HOTAIRM1 has demonstrated involvement in biological and tumor characteristics and is upregulated in various cancers including glioma [24,32], acute myeloid leukemia [33,35], osteosarcoma [36], endometrial cancer [37], thyroid cancer [38,39], non-small cell lung cancer [41,42], ORAL squamous cell carcinoma [43], prostate cancer [44], pancreatic ductal adenocarcinoma [45,46], ovarian cancer [47] (Table 1). On the other hand, expression of this lncRNA has been down-regulated in papillary thyroid cancer [40], ovarian cancer [48], head and neck tumor [49], hepatocellular carcinoma [50], lung adenocarcinoma [51], gastric cancer [52-53], colorectal cancer [54,55] (Table 2).
HOTAIRM1 expression in glioma: Glioma is the most lethal primary central nervous system tumor. In recent years, several studies have demonstrated that HOTAIRM1 expression was significantly increased in GBM tissues and GBM cell lines in comparison with normal counterparts [24-29]. A recent study described that HOTAIRM1 was abnormally up-regulated in GBM tissues and cells, and this up-regulation was correlated with grade malignancy in glioma patients. The high expression of HOTAIRM1 was modulated by METTL3-dependent m6A modification [30]. Xie et al. found that HOTAIRM was transcriptionally activated by SNAI2, and high HOTAIRM1 expression was evidently associated with poorer prognosis in glioma [31]. Additionally, Ahmadov et al. showed HOTAIRM1 as a lncRNA whose up-regulation was significantly associated with shorter survival of glioblastoma patients [32].
Table 1: HOTAIRM1 expression in various cancers.
| Cancer types | Expression | Functions | Relatedgenes | Role | Refs |
|---|---|---|---|---|---|
| Glioma | Up | migration, invasion, VM formation,proliferation, stemness, radiosensitivity | IGFBP2, FUS, HOXAs, miR-133b-3p,miR-137, miR-153-5p, TGM2 | Oncogene | 24-32 |
| AML | Up | autophagy, proliferation, apoptosis,cellcycle, differentiation | EGR1, miR-152-3p,miR-148b | Oncogene | 33-35 |
| OS | Up | proliferation, apoptosis | miR-664b-3p | Oncogene | 36 |
| EC | Up | proliferation, migration, invasion, EMT | HOXA1 | Oncogene | 37 |
| TC | Up | proliferation, apoptosis, migration, invasion | ILF3, pri-miR-144, miR‑148a | Oncogene | 38-39 |
| NSCLC | Up | proliferation, apoptosis, migration,invasion, glycolysis metabolism | miR-498 | Oncogene | 41-42 |
| OSCC | Up | Proliferation, cell cycle | PCNA, CyclinD1, p53, p21, CDK4, CDK6 | Oncogene | 43 |
| PCa | Up | proliferation, apoptosis | β-catenin | Oncogene | 44 |
| PDAC | Up | poliferation, apoptosis, cell cycle, migration | CDK1,Cyclin D1, p21, Bax,Bad, Bcl-2 | Oncogene | 45-46 |
| OC | Up | Proliferation, Apoptosis | MMP2 | Oncogene | 47 |
HOTAIRM1 expression in acute myeloid leukemia: Acute myeloid leukemia (AML) is a heterogeneous disease characterized by genetic abnormalities and epigenetic changes. Jing et al. reported that HOTAIRM1 expression was upregulated in 14 NPM1-mutated AML samples compared with corresponding NPM1-unmutated AML samples. Moreover, Kaplan-Meier survival analysis of AML patients revealed a significantly shorter overall survival time in the cohort with high HOTAIRM1 expres sion [33]. Another study revealed that HOTAIRM1 was up-regulated in APL cells upon exposure to ATRA and played a critical role in myeloid differentiation. Importantly, PU.1 could transactive HOTAIRM1 through the +1100 region in the HOTAIRM1 promoter [34]. Furthermore, Hu et al. showed that HOTAIRM1 expression was higher in AML patients than healthy controls. Functional analysis revealed that depletion of HOTAIRM1 suppressed proliferation and induced apoptosis in AML cells, suggesting that HOTAIRM1 played an oncogenic role in AML [35].
Table 2: HOTAIRM1 expression in various cancers.
| Cancer types | Expression | Functions | Relatedgenes | Role | Refs |
|---|---|---|---|---|---|
| PTC | Down | proliferation, migration, invasion | miR-107 |
Anti-cancer | 40 |
| OC | Down | proliferation invasion, apoptosis | miR-106a-5p | Anti-cancer | 48 |
| HNT | Down | proliferation, apoptosis, migration,invasion | miR-148a | Anti-cancer | 49 |
| HCC | Down | proliferation, apoptosis | β-catenin | Anti-cancer | 50 |
| ADC | Down | cell cycle, proliferation, invasion | miR-498 | Anti-cancer | 51 |
| GC | Down | proliferation, migration, apoptosis | miR-29b-1-5p, miR-17-5p | Anti-cancer | 52-53 |
| CRC | Down | invasion, migration, multi-drugresistance | miR-17-5p | Anti-cancer | 54-55 |
HOTAIRM1 expression in endometrial cancer: Endometrial cancer (EC) is one of the three most common malignancies of the female reproductive system. Li et al. discovered extreme upregulation of HOTAIRM1 in type I EC tissues than that in normal endometrium tissues. HOTAIRM1 expression was also linked to the clinicopathological characteristics of type I EC patients, and analysis revealed that upregulated HOTAIRM1 was closely correlated with advanced FIGO stage and lymph node metastasis. Concerning biological function, knockdown of HOTAIRM1 could inhibit tumor cell proliferation, migration, invasion and epithelial–mesenchymal transition (EMT) of type I EC cells. In addition, silencing HOTAIRM1 suppressed the growth of tumors in vivo [37].
HOTAIRM1 expression in thyroid cancer: Thyroid cancer (TC) is categorised into three broad histological categories: (1) differentiated thyroid cancer, which includes papillary, follicular, and oncocytic thyroid carcinoma; (2) medullary thyroid cancer, which is sometimes associated with the multiple endocrine neoplasia type 2 syndromes; and (3) anaplastic thyroid cancer, which often arises from differentiated thyroid cancer and has a high mortality. Zhang et al. found that HOTAIRM1, whose encoding gene was amplified and expression was upregulated in ATC compared with papillary thyroid cancer and normal thyroid. Increased genomic copy number and expression of HOTAIRM1 were both correlated with poor survival of ATC patients [38]. Li et al. pronounced that upregulated expression of HOTAIRM1 was evident in TC cells and tissues, and was associated with TNM stage and lymph node metastasis [39]. Another study reported that HOTAIRM1 was significantly downregulated in PTC tissues and low HOTAIRM1 expression levels were associated with lymph node metastasis and advanced TNM stage [40].
HOTAIRM1 expression in non-small cell lung cancer: Lung cancer is the leading cause of cancer deaths worldwide, and non-small cell lung cancer (NSCLC) accounts for nearly 85%. Chen et al. found that HOTAIRM1 expression was obviously upregulated in NSCLC tissues as compared to control groups [41]. Another study showed that a significant association between the expression of HOTAIRM1 and histopathological differentiation, tumor size, tumor/node/metastasis (TNM) stage, and Ki-67 of NSCLC patients. In addition, the relative expression of HOTAIRM1 in NSCLC tissues was significantly higher when compared with the individual patients’ matched paracancerous tissues. Patients in the group with low expression levels of HOTAIRM1 had a longer overall survival than those in the group with high expression levels for NSCLC [42].
HOTAIRM1 expression in ORAL squamous cell carcinoma: ORAL squamous cell carcinoma (OSCC) is a malignant tumor with the highest incidence among tumors involving the oral cavity maxillofacial region, and is notorious for its high recurrence and metastasis potential. Yu et al. reported that HOTAIRM1 as a novel significantly upregulated lncRNA in OSCC, which was strongly associated with unfavorable prognosis of OSCC. Systematic bioinformatics analyses demonstrated that HOTAIRM1 was closely related to tumor stage, overall survival, genome instability, the tumor cell stemness, the tumor microenvironment, and immunocyte infiltration [43].
HOTAIRM1 expression in prostate cancer: Prostate cancer (PCa) is a leading cause of cancer-associated mortality among men, specifically in Western countries; the lowest rates of this cancer are seen in Africa and Asia. Wang et al. corroborated that HOTAIRM1 was significantly overexpressed in prostate cancer cells. In vitro, silencing HOTAIRM1 attenuated tumor cell proliferation but gave rise to apoptosis by regulating the expression of proteins related to the cell apoptosis. Knocking down HOTAIRM1 could restrain the Wnt pathway in prostate cancer cells, consequently suppressing the malignant biological behaviors of tumor cells [44].
HOTAIRM1 expression in pancreatic ductal adenocarcinoma: Pancreatic ductal adenocarcinoma (PDAC) is the major form of pancreatic cancer, which derives from the epithelial cells of pancreatic duct. HOTAIRM1 expression was anomalously upregulated in 47 PDAC tissues and 5 cell lines compared with the samples in the control group [45]. Similarly, Zhou et al. reported that the expression level of HOTAIRM1 was upregulated in 12 PDAC tissues samples compared with matched adjacent non-tumor samples [46].
HOTAIRM1 expression in ovarian cancer: Ovarian cancer (OC) ranks as the eighth most prevalent form of cancer in women across the globe and stands as the third most frequent gynecological cancer, following cervical and endometrial cancers. Ye et al. found that HOTAIRM1 was overexpressed in SKOV3 cells. Silencing of HOTAIRM1 alleviated cell proliferation, and increased cell apoptosis [47]. Notably, Chao et al. identified that HOTAIRM1 expression was lowered in OC tumor tissues and cells. In that study, decreased HOTAIRM1 expression was associated with advanced FIGO stages and lymphatic metastasis. Up-regulation of HOTAIRM1 suppressed OC cell proliferation and invasion, and promoted apoptosis. Also, HOTAIRM1 slowed OC tumor growth in vivo [48].
HOTAIRM1 expression in head and neck tumor: Head and neck tumor (HNT) is one of the most common diagnosed malignancies worldwide, with over 600,000 new cases have reported each year. Zheng et al. discovered that the expression level of HOTAIRM1 in 43 Head carcinoma tissues and 41 Neck carcinoma tissues was downregulated compared to the corresponding adjacent normal tissues. In addition, there was no significant association between HOTAIRM1 level and age, gender, or tu mor location, while patients with high expression of HOTAIRM1 were more potential to get an advanced TNM stage [49].
HOTAIRM1 expression in hepatocellular carcinoma: Hepatocellular carcinoma (HCC) is ranked as the sixth most common neoplasm and the third leading cause of cancer death, and is an important medical problem. Zhang et al. discovered that lower lncRNA HOTAIRM1 expressions were observed in the HCC tissues than those of the paracancerous tissues. Moreover, ROC curve indicated a high sensitivity and specificity of HOTAIRM1 for HCC. Progression-free survival in HCC patients was correlated with tumor size and HOTAIRM1 expression, whereas not correlated with age, sex, GGT, AFP, Child-Pugh grade, HBsAg, cirrhosis, number of tumors, micro-vessel metastasis, tumor differentiation, and TNM stage of HCC [50].
HOTAIRM1 expression in lung adenocarcinoma: Lung adenocarcinoma (ADC) is a major form of lung cancer, which is a main cause of global cancer-related death in male and female patients. Chen et al. showed that HOTAIRM1 expression was remarkably reduced in ADC tissues compared with those in normal lung tissues. An overt association was observed between the knockdown of HOTAIRM1 expression and clinical stage, and lymph node metastasis and tumor size. Additionally, the knockdown of HOTAIRM1 was linked to worse overall survival of ADC patients, as indicated by Kaplan-Meier analysis [51].
HOTAIRM1 expression in gastric cancer: Gastric cancer (GC) is one of the most common cancers in the world. In China, it is the second most common cancer and the third leading cause of cancer-related deaths. Xu et al. found that HOTAIRM1 expression was significantly down-regulated in GC tissues, and the low expression of HOTAIRM1 was associated with the shorter survival rate of GC patients [52]. These results are similar to the observation reported by Lu et al., which indicated that HOTAIRM1 expression was obviously lower in 20 GC tissues and cell lines. More notably, HOTAIRM1 expression was also linked to the clinicopathological characteristics of GC patients, and analysis revealed that downregulated HOTAIRM1 was closely correlated with advanced TNM stage and lymph node metastasis [53].
HOTAIRM1 expression in colorectal cancer: Colorectal cancer (CRC) is one of the most common cancers worldwide, especially in Western countries. Wan et al. discovered that HOTAIRM1 expression was downregulated in CRC tissues compared with paired normal tissues, and in the matched group, there were lower plasma HOTAIRM1 levels in CRC patients [54]. A similar situation was found in Ren et al., which showed that HOTAIRM1 is downregulated in CRC tissues and cell line, and even lower in 5-FU resistant CRC tissues and cell lines [55].
Mechanism of HOTAIRM1 regulations: LncRNAs interact with DNA, RNA or proteins as molecular sponges, scaffolds and activators to play important regulatory roles in a variety of biological processes ranging from gene regulation, cellular differentiation to human diseases, especially in cancers [56-60]. HOTAIRM1 has been shown to participate in both physiological and pathological processes. Decades of study have identified some molecular functions, which can be classified into three main pathways.
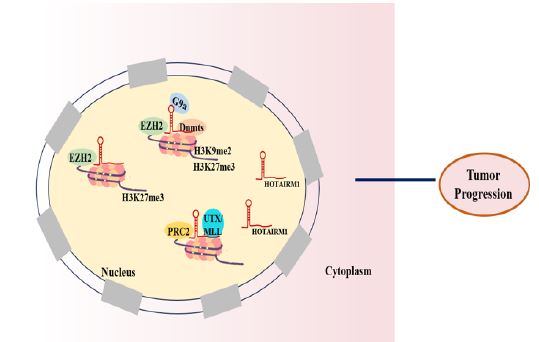
Interaction with DNA: HOTAIRM1 is involved in the methylation modification of a series of tumor-related genes and in histone modification (Figure 2). HOTAIRM1 could regulate gene expression by interacting with Polycomb Repressive Complex 2 (PRC2), which is composed of EZH2, SUZ12, and EED. It can catalyze the dimethylation and trimethylation of the lysine residue 27 of histone 3 (H3K27me3), thereby regulating its gene expression level. Li et al. found that HOTAIRM1 activated transcription of the HOXA1 gene through the decrease of histone H3K9me2, H3K27me3 and DNA methylation, which are epigenetic markers associated with gene silencing. While HOTAIRM1 interacting with G9a, EZH2 and DNA methyltransferases Dnmts, occlude them from the promoter of HOXA1 gene, hence reducing their enrichment [61]. Kim et al. revealed that HOTAIRM1 directly interacted with EZH2, the H3K27me3 specific methyltransferases, hindered the deposition of H3K27me3 marks at the putative HOXA1 promoter, resulting in the higher expression of HOXA1 in ER+ breast cancer cells [62]. Furthermore, HOTAIRM1 bond to PRC2 and histone demethylase UTX/MLL to regulate chromatin conformation and then affected HOXA gene cluster transcriptional activity [63].
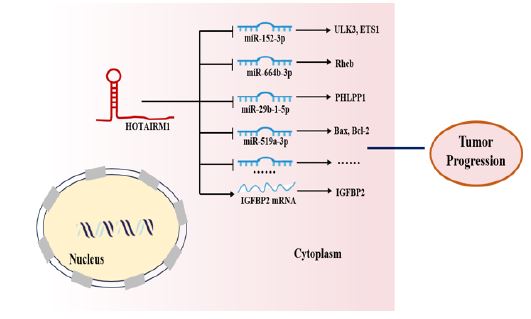
Interaction with RNA: In addition to DNA, HOTAIRM1 can interact with RNA to execute diverse molecular functions. In recent years, the importance of competitive endogenous RNA (ceRNA) networks in the formation and development of cancer has emerged [64,65]. MicroRNAs are important regulators in the ceRNA network, which negatively regulates mRNA expression. The most common mechanism of which is the interaction between miRNAs and the mRNA 3’-UTRs of the target genes, which leads to mRNA demethylation and destabilization [66,67]. HOTAIRM1 competitively binds miRNA through the ceRNA mechanism, increases the expression of downstream mRNA, and regulates tumor progression (Figure 3). Investigations had reported that HOTAIRM1, which worked as a ceRNA for miR-152-3p to modulate ULK3, regulated leukemic cell function, such as cell autophagy, proliferation and apoptosis [33]. Yu et al. found that HOTAIRM1 competitively bond miR-664b-3p and upregulated the expression of Rheb to promote the mTOR pathway-mediated Warburg effect in OS [36]. HOTAIRM1 could competitively bind miR-152-3p and inhibited its expression, increased that of ETS1 mRNA, and promoted osteogenic differentiation of human bone marrow‑derived mesenchymal stem cells [68]. Similarly, HOTAIRM1 acted as a sponge for miR-29b-1-5p and PHLPP1 was regulated by the HOTAIRM1/miR-29b-1-5p axis in GC cells [52]. Furthermore, Wang et al. reported that HOTAIRM1/miR-519a-3p axis played key roles in proliferation, apoptosis, inflammation, and oxidative stress of neuroblastoma cells treated with MPP+ [69]. HOTAIRM1 has been reported to directly regulate mRNA metabolism. HOTAIRM1 bond to IGFBP2 mRNA, up-regulated the expression of IGFBP2, promoted cell proliferation, migration, invasion, and VM formation in glioma [24].
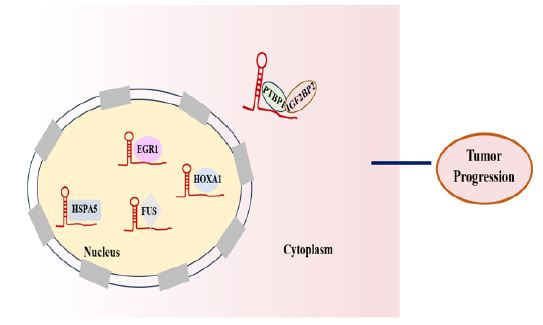
Interaction with protein: Recent studies have shown that many lncRNAs directly bind proteins to participate in molecular regulation, and many proteins have been shown to interact with HOTAIRM1 (Figure 4). For instance, HOTAIRM1 interacted with HSPA5, regulated HSPA5 expression in a transcription‑dependent manner and its effect on proliferation was HSPA5 partially dependent [71]. Han et al. confirmed that HOTAIRM1 physically bond to PTBP1 and IGF2BP2, strengthened the interaction between PTBP1 and IGF2BP2, and recruited them to bind to SHMT2 mRNA, in turn led to an increase in the protein level of SHMT2 by enhancing the stability of its mRNA, resulting in the induction of mitochondrial activity and the malignant progression of glioma [72]. Liu et al. demonstrated that HOTAIRM1 could bind to FUS, an RNA-binding protein (RBP), and thus regulated E2F7, which promoted the proliferation, migration, and invasion of tMSCs transformed by glioma stem cells in the glioma microenvironment [25]. In addition, Chen et al. showed that HOTAIRM1 regulated PD-L1 expression in lung alveolar epithelial cells by binding to a key transcription, HOXA1 [73]. Jing et al. showed that the association of HOTAIRM1 with both EGR1 and MDM2, HOTAIRM1 acts as a scaffold to recruit MDM2 to EGR1 to promote leukemia cell autophagy and proliferation [33].
HOTAIRM1 regulates various hallmarks of cancer: Cancer is a complex disease involving multiple gene mutations, including epigenetic changes, chromosomal translocations, deletions, and increases. The genome encodes lncRNAs, but most are not translated into proteins. Although there is no translation, lncRNAs have crucial physiological functions in various cell [73,74]. Recent studies revealed that HOTAIRM1 is associated with hallmarks of cancer, including proliferation, cell death, invasion and metastasis, metabolic reprogramming, and angiogenesis, by regulating various pathways (Figure 5).
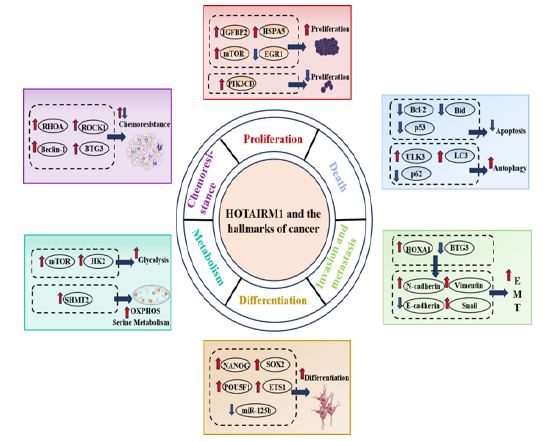
Proliferation: Abnormal, uncontrolled cell growth due to the dysregulation of cell proliferation is the most fundamental cause of tumorigenesis [75]. Aberrant HOTAIRM1 expression has been observed in various tumors and is closely linked with tumor cell proliferation. For example, Wu et al. reported that HOTAIRM1 presented up-regulation in glioma tissues and cells and overexpression of HOTAIRM1 facilitated glioma cell proliferation [30]. Zhou et al. delineated a pivotal functional interaction among HOXA4, HOTAIRM1, and HSPA5 as a novel regulatory circuit for modulating HUVEC proliferation [70]. HOTAIRM1 could promote the growth and proliferation of OS cells by sponging miR-664b-3p, thereby activating the mTOR pathway [36]. On the other hand, HOTAIRM1 also could reduce the proliferative ability of granulosa cells [76]. Additionally, Jing et al. indicated an important role of HOTAIRM1 in promoting proliferation in NPM1-mutated AML cells [33].
Death: Apoptosis is a programmed cell death controlled by a signaling cascade to maintain a stable internal environment. The elimination of cancer cells by apoptosis has been a key cue in clinical cancer treatment. Dahariya et al. found that HOTAIRM1 acted a decoy for miR-125b to control cell apoptosis to support terminal maturation of megakaryocytes [77]. HOTAIRM1 also could induce Jurkat cell apoptosis through KIT/AKT signaling pathway [78]. Liu et al. revealed that the loss of HOTAIRM1 function led to aberrant increases in the apoptosis of chondrocytes [79]. Furthermore, Ye et al. showed that HOTAIRM1 knockdown significantly increased expression of pro-apoptotic agents, such as Bad and Bax, while it decreased expressions of Bid and Bcl-2 (anti-apoptotic agents) in ovarian cancer cells [47].
Autophagy is an intracellular self-destructive form of cell death that transfers cytoplasmic proteins or organelles to the lysosome to fulfill the metabolic and self-renewal needs of organelles and the cell itself. Previous studies have reported that HOTAIRM1 could activate autophagy by promoting the ULK3 expression, leading to the increase in autophagy-related proteins microtubule-associated protein LC3 II as well as the decrease in the autophagy substrate p62 [33].
Invasion and metastasis: Metastasis is a complex process that includes EMT, invasion, intravasation, cell survival in circulation, extravasation, and metastatic colonization. EMT is the first, initiative event in cancer metastasis in which epithelial cells gained mesenchymal characteristics such as decreased intercellular adhesion and increased motility, while losing epithelial characteristics. HOTAIRM1 could promote type I EC cells’ EMT and metastasis potential by upregulating HOXA1 expression, leading to the decrease in the expression of epithelial marker E-cadherin, and an increase in mesenchymal markers such as N-cadherin [37]. Ren et al. investigated the effects of HOTAIRM1 dysregulation in 5-FU resistant CRC cells and found that its overexpression could reduce cell invasion and migration by EMT assays [55].
Differentiation: Cell differentiation is the formation of different tissues, the pre - and post-differentiation cells do not belong to the same type. Carcinogenesis is an abnormal differentiation of cells. Tollis et al. used a model system that recapitulates spinal MN (spMN) differentiation to show nHOTAIRM1 intervened in the binary cell fate decision between MNs and interneurons, acting as a pro-MN factor [80]. A recent study indicated that HOTAIRM1 participated in proper progression through the early stages of neuronal differentiation, HOTAIRM1 affected the NANOG/POU5F1/SOX2 core pluripotency network maintaining an undifferentiated cell state [81]. Wang et al. identified that HOTAIRM1 as a novel regulator of osteogenic differentiation of BMSCs by the regulation of miR-152-3p/ETS1 axis, uncovering HOTAIRM1 as a promising therapeutic strategy for osteoporosis [68]. Furthermore, Dahariya et al. revealed that the regulatory role of HOTAIRM1 in p53-mediated regulation of cyclin D1 during megakaryocytopoiesis is to promote MK maturation by decoying miR-125b [77].
Metabolism: Metabolic alteration is a characteristic of tumor cells crucial for supporting their rapid cell growth. Unlike normal cells, which mainly depend on glycolysis followed by oxidative phosphorylation, tumor cells prefer inefficient aerobic glycolysis with a significantly higher turnover rate compared to normal cells even under adequate oxygen availability. This phenomenon is known as the Warburg effect. HOTAIRM1 dramatically enhanced the Warburg effect in Osteosarcoma cells via the miR-664b-3p/Rheb/mTOR axis [36]. A recent study showed that HOTAIRM1 knockdown affected the Warburg effect in NSCLC. In this study, Glucose uptake and lactate production assays showed that HOTAIRM1 knockdown repressed the glucose uptake and lactate production. Western blot results explained that the protein level of HK2 was dramatically repressed by HOTAIRM1 depletion [41]. Han et al. found that HOTAIRM1 led to an increase in the protein level of SHMT2 by enhancing the stability of its mRNA, resulting in the induction of oxidative phosphorylation and serine metabolism in mitochondrial activity [71].
Chemoresistance: Drug resistance is a major obstacle to the effective treatment of cancer patients. Recent studies have shown that HOTAIRM1 has been implicated in lenvatinib resistance of Hepatocellular carcinoma cancer cells, and HOTAIRM1 expression induced lenvatinib resistance via downregulating miR-34a expression and upregulating the expression of Beclin-1 [82]. Liang et al. determined that HOTAIRM1 bound to the transcriptional inhibitory region of ARHGAP18 and repressed the expression of ARHGAP18, which led to the increase of RHOA/ROCK1 signaling pathway and promoted GC resistance of leukemia cells [83]. In addition, Chen et al. showed that knockdown of HOTAIRM1 enhanced Ara-C cytotoxicity through regulating the Wnt/β-catenin/PFKP signaling pathway [84]. HOTAIRM1 might act as a tumor-suppressor in 5-FU resistant CRC cells in vitro and in vivo through downregulating miR-17-5p/BTG3 pathway and inhibiting multi-drug resistance [55].
Conclusions
Cancer poses a severe threat to human health globally. Although the overall mortality rate of cancer has declined in recent years, its early diagnosis and treatment still face major challenges. Many patients with malignant tumors are diagnosed at a late stage and have a poor prognosis. Therefore, it is imperative to search for novel biomarkers and explore the various molecular mechanisms for early diagnosis and treatment of cancer.
Numerous studies have shown that lncRNAs are abnormally expressed in many diseases and can act as tumor suppressors or oncogenes. HOTAIRM1 is abnormally expressed in various tumor tissues and cells, and can be an independent predictor of poor prognosis in diverse carcinomas. On the hand, HOTAIRM1 overexpression promotes tumor cell growth in many neoplasms, including Glioma, AML, OS, EC, TC, NSCLC, OSCC, PCa, PDAC and OC. On the other hand, forced expression of HOTAIRM1 suppresses cancer cell proliferation in PTC, OC, HNT, HCC, ADC, GC and CRC. Moreover, abnormal expression of HOTAIRM1 in tumors is closely related to a variety of clinical and pathological features such as age, tumor size, vascular invasion, distant metastasis, overall survival, and neoplasm recurrence.
In the review, as described in Figure 2-4, we summarized the specific mechanism by which HOTAIRM1 acts as a tumor regulator. Firstly, as described in Figure 2, HOTAIRM1 can induce H3K27me3 by interacting with EZH2/PRC2 and can also involve in chromatin remodeling by directing the recruitment of chromatin modifiers to target gene site. Second, as described in Figure 3, HOTAIRM1 acts as a molecular sponge for miRNAs and plays its role through the HOTAIRM1-miRNA-mRNA axis. In addition, as described in Figure 4, HOTAIRM1 can bind to nuclear/cytoplasmic functional protein to regulate its expression to play an important role in tumor progression. Indeed, the other possible molecular mechanisms of the biological effects of HOTAIRM1 merits further exploration.
HOTAIRM1 has emerged as a potential tumor regulator that could affect various hallmarks of cancer including cell proliferation, cell death, invasion and metastasis, cell differentiation, cell metabolism and chemoresistance (Figure 5). However, in some cases, for example in drug resistance, the role of HOTAIRM1 is still unclear, as current studies provide paradoxical results that require further detailed investigation. Liang et al. revealed that HOTAIRM1 promoted GC resistance through anti-apoptosis of leukemia cells. Notably, Ren et al. reported that HOTAIRM1 downregulated miR-17-5p/BTG3 pathway and inhibited multi-drug resistance. At present, while more detailed studies are still needed to investigate whether HOTAIRM1 could regulate other hallmarks of cancer, such as avoiding immune destruction, genome instability and mutation, non-mutational epigenetic reprogramming, unlocking phenotypic plasticity and polymorphic microbiomes.
Declarations
Author contributions: Conceptualization, Y.J.; writing—original draft preparation, Y.J., S.W. and Y.L.; writing-review and editing, Y.J., Y.G., X.Q., and Y.L.; supervision, Y.L.; project administration, Y.J.; funding acquisition, Y.L.; All authors have read and agreed to the published version of the manuscript.
Funding: This work was supported by National Key Research and Development Program of China (2018YFE0207300), the National High Level Hospital Clinical Research Funding (2022-PUMCH-B-124) and supported by Beijing Natural Science Foundation (M23008).
Conflicts of interest: The authors declare no conflict of interest.
References
- Vaghari-Tabari M, Ferns GA, Qujeq D, Andevari AN, Sabahi Z, MoeinZ S. Signaling, metabolism, and cancer: An important relationship for therapeutic intervention. J Cell Physiol. 2021; 236: 5512-5532.
- Giunta S. Decoding human cancer with whole genome sequencing: a review of PCAWG Project studies published in February 2020. Cancer Metastasis Rev. 2021; 40: 909-924.
- Chi Y, Wang D, Wang J, Yu W, Yang J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells. 2019; 8: 1015.
- Ramezankhani R, Solhi R, Es HA, Vosough M, Hassan M. Novel molecular targets in gastric adenocarcinoma. Pharmacol Ther. 2021; 220: 107714.
- Guo N, Liu JB, Li W, Ma YS, Fu D. The power and the promise of CRISPR/Cas9 genome editing for clinical application with gene therapy. J Adv Res. 2022; 40: 135-152.
- Traba J, Sack MN, Waldmann TA, Anton OM. Immunometabolism at the Nexus of Cancer Therapeutic Efficacy and Resistance. Front Immunol. 2021; 12: 657293.
- Ghafouri-Fard S, Khoshbakht T, Taheri M, Hajiesmaeili M. Long intergenic non-protein coding RNA 460: Review of its role in carcinogenesis. Pathol Res Pract. 2021; 225: 153556.
- Hovhannisyan H, Gabaldón T. The long non-coding RNA landscape of Candida yeast pathogens. Nat Commun. 2021; 12: 7317.
- Kovalenko TF, Yadav B, Anufrieva KS, Rubtsov YP, Zatsepin TS, Shcherbinina EY, Solyus EM, Staroverov DB, Larionova TD, Latyshev YA, et al. Functions of long non-coding RNA ROR in patient-derived glioblastoma cells. Biochimie. 2022; 200: 131-139.
- Statello L, Guo CJ, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021; 22: 96-118.
- Herman AB, Tsitsipatis D, Gorospe M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol Cell. 2022; 82: 2252-2266.
- Sun KK, Zu C, Wu XY, Wang QH, Hua P, Zhang YF, Shen XJ, Wu YY. Identification of lncRNA and mRNA regulatory networks associated with gastric cancer progression. Front Oncol. 2023; 13: 1140460.
- Pierce JB, Zhou H, Simion V, Feinberg MW. Long Noncoding RNAs as Therapeutic Targets. Adv Exp Med Biol. 2022; 1363: 161-175.
- Sun W, Xu J, Wang L, Jiang Y, Cui J, Su X, Yang F, Tian L, Si Z, Xing Y. Non-coding RNAs in cancer therapy-induced cardiotoxicity: Mechanisms, biomarkers, and treatments. Front Cardiovasc Med. 2022; 9: 946137.
- Zhu Y, Ren J, Wu X, Zhang Y, Wang Y, Xu J, Tan Q, Jiang Y, Li Y. lncRNA ENST00000422059 promotes cell proliferation and inhibits cell apoptosis in breast cancer by regulating the miR-145-5p/KLF5 axis. Acta Biochim Biophys Sin (Shanghai). 2023; 55: 1892-1901.
- Zhang X, Lian Z, Padden C, Gerstein MB, Rozowsky J, Snyder M, Gingeras TR, Kapranov P, Weissman SM, Newburger PE. A myelopoiesis-associated regulatory intergenic noncoding RNA transcript within the human HOXA cluster. Blood. 2009; 113:2526-34.
- Zhang X, Weissman SM, Newburger PE. Long intergenic non-coding RNA HOTAIRM1 regulates cell cycle progression during myeloid maturation in NB4 human promyelocytic leukemia cells. RNA Biol. 2014; 11: 777-87.
- Chen R, Zhou D, Chen Y, Chen M, Shuai Z. Understanding the role of exosomal lncRNAs in rheumatic diseases: a review. PeerJ. 2023; 11: 16434.
- Ramos TAR, Urquiza-Zurich S, Kim SY, Gillette TG, Hill JA, Lavandero S, Rêgo TG, Maracaja-Coutinho V. Single-cell transcriptional landscape of long non-coding RNAs orchestrating mouse heart development. Cell Death Dis. 2023; 14: 841.
- Hu C, Dai Q, Zhang R, Yang H, Wang M, Gu K, Yang J, Meng W, Chen P, Xu M. Case Report: Identification of a novel LYN: LINC01900 transcript with promyelocytic phenotype and TP53 mutation in acute myeloid leukemia. Front Oncol. 2023; 13: 1322403.
- Wang D, Zhao X, Li S, Guo H, Li S, Yu D. The impact of LncRNA-SOX2-OT/let-7c-3p/SKP2 Axis on head and neck squamous cell carcinoma progression: Insights from bioinformatics analysis and experimental validation. Cell Signal. 2023; 111018.
- Yu M, He X, Liu T, Li J. lncRNA GPRC5D-AS1 as a ceRNA inhibits skeletal muscle aging by regulating miR-520d-5p. Aging (Albany NY). 2023; 15: 13980-13997.
- Zhang X, Zhong Y, Liu L, Jia C, Cai H, Yang J, Wu B, Lv Z. Fasting regulates mitochondrial function through lncRNA PRKCQ-AS1-mediated IGF2BPs in papillary thyroid carcinoma. Cell Death Dis. 2023; 14: 827.
- Wu Z, Lin Y, Wei N. N6-methyladenosine-modified HOTAIRM1 promotes vasculogenic mimicry formation in glioma. Cancer Sci. 2023; 114: 129-141.
- Liu L, Zhou Y, Dong X, Li S, Cheng S, Li H, Li Y, Yuan J, Wang L, Dong J. HOTAIRM1 Maintained the Malignant Phenotype of tMSCs Transformed by GSCs via E2F7 by Binding to FUS. J Oncol. 2022; 2022: 7734413.
- Shi T, Guo D, Xu H, Su G, Chen J, Zhao Z, Shi J, Wedemeyer M, Attenello F, Zhang L, et al. HOTAIRM1, an enhancer lncRNA, promotes glioma proliferation by regulating long-range chromatin interactions within HOXA cluster genes. Mol Biol Rep. 2020; 47: 2723-2733.
- Wang H, Li H, Jiang Q, Dong X, Li S, Cheng S, Shi J, Liu L, Qian Z, Dong J. HOTAIRM1 Promotes Malignant Progression of Transformed Fibroblasts in Glioma Stem-Like Cells Remodeled Microenvironment via Regulating miR-133b-3p/TGFβ Axis. Front Oncol. 2021; 11: 603128.
- Xia H, Liu Y, Wang Z, Zhang W, Qi M, Qi B, Jiang X. Long Noncoding RNA HOTAIRM1 Maintains Tumorigenicity of Glioblastoma Stem-Like Cells Through Regulation of HOX Gene Expression. Neurotherapeutics. 2020; 17: 754-764.
- Hao Y, Li X, Chen H, Huo H, Liu Z, Chai E. Over-expression of long noncoding RNA HOTAIRM1 promotes cell proliferation and invasion in human glioblastoma by up-regulating SP1 via sponging miR-137. Neuroreport. 2020; 31: 109-117.
- Wu Z, Wei N. METTL3-mediated HOTAIRM1 promotes vasculogenic mimicry icontributionsn glioma via regulating IGFBP2 expression. J Transl Med. 2023; 21: 855.
- Xie P, Li X, Chen R, Liu Y, Liu DC, Liu W, Cui G, Xu J. Upregulation of HOTAIRM1 increases migration and invasion by glioblastoma cells. Aging (Albany NY). 2020; 13: 2348-2364.
- Ahmadov U, Picard D, Bartl J, Silginer M, Trajkovic-Arsic M, Qin N, Blümel L, Wolter M, Lim JKM, Pauck D, et al. The long non-coding RNA HOTAIRM1 promotes tumor aggressiveness and radiotherapy resistance in glioblastoma. Cell Death Dis. 2021; 12: 885.
- ing Y, Jiang X, Lei L, Peng M, Ren J, Xiao Q, Tao Y, Tao Y, Huang J, Wang L, et al. Mutant NPM1-regulated lncRNA HOTAIRM1 promotes leukemia cell autophagy and proliferation by targeting EGR1 and ULK3. J Exp Clin Cancer Res. 2021; 40: 312.
- Wei S, Zhao M, Wang X, Li Y, Wang K. PU.1 controls the expression of long noncoding RNA HOTAIRM1 during granulocytic differentiation. J Hematol Oncol. 2016; 9: 44.
- Hu N, Chen L, Li Q, Zhao H. LncRNA HOTAIRM1 is involved in the progression of acute myeloid leukemia through targeting miR-148b. RSC Adv. 2019; 9: 10352-10359.
- Yu X, Duan W, Wu F, Yang D, Wang X, Wu J, Zhou D, Shen Y. LncRNA-HOTAIRM1 promotes aerobic glycolysis and proliferation in osteosarcoma via the miR-664b-3p/Rheb/mTOR pathway. Cancer Sci. 2023; 114: 3537-3552.
- Li X, Pang L, Yang Z, Liu J, Li W, Wang D. LncRNA HOTAIRM1/HOXA1 Axis Promotes Cell Proliferation, Migration And Invasion In Endometrial Cancer. Onco Targets Ther. 2019; 12: 10997-11015.
- Zhang L, Zhang J, Li S, Zhang Y, Liu Y, Dong J, Zhao W, Yu B, Wang H, Liu J. Genomic amplification of long noncoding RNA HOTAIRM1 drives anaplastic thyroid cancer progression via repressing miR-144 biogenesis. RNA Biol. 2021; 18: 547-562.
- Li C, Chen X, Liu T, Chen G. lncRNA HOTAIRM1 regulates cell proliferation and the metastasis of thyroid cancer by targeting Wnt10b. Oncol Rep. 2021; 45: 1083-1093.
- Li D, Chai L, Yu X, Song Y, Zhu X, Fan S, Jiang W, Qiao T, Tong J, Liu S, et al. The HOTAIRM1/miR-107/TDG axis regulates papillary thyroid cancer cell proliferation and invasion. Cell Death Dis. 2020; 11: 227.
- Chen D, Li Y, Wang Y, Xu J. LncRNA HOTAIRM1 knockdown inhibits cell glycolysis metabolism and tumor progression by miR-498/ABCE1 axis in non-small cell lung cancer. Genes Genomics. 2021; 43: 183-194.
- Xiong F, Yin H, Zhang H, Zhu C, Zhang B, Chen S, Ling C, Chen X. Clinicopathologic Features and the Prognostic Implications of Long Noncoding RNA HOTAIRM1 in Non-Small Cell Lung Cancer. Genet Test Mol Biomarkers. 2020; 24: 47-53.
- Yu Y, Niu J, Zhang X, Wang X, Song H, Liu Y, Jiao X, Chen F. Identification and Validation of HOTAIRM1 as a Novel Biomarker for Oral Squamous Cell Carcinoma. Front Bioeng Biotechnol. 2022; 9: 798584.
- Wang L, Wang L, Wang Q, Yosefi B, Wei S, Wang X, Shen D. The function of long noncoding RNA HOTAIRM1 in the progression of prostate cancer cells. Andrologia. 2021; 53: 13897.
- Luo Y, He Y, Ye X, Song J, Wang Q, Li Y, Xie X. High Expression of Long Noncoding RNA HOTAIRM1 is Associated with the Proliferation and Migration in Pancreatic Ductal Adenocarcinoma. Pathol Oncol Res. 2019; 25: 1567-1577.
- Zhou Y, Gong B, Jiang ZL, Zhong S, Liu XC, Dong K, Wu HS, Yang HJ, Zhu SK. Microarray expression profile analysis of long non-coding RNAs in pancreatic ductal adenocarcinoma. Int J Oncol. 2016; 48: 670-80.
- Ye L, Meng X, Xiang R, Li W, Wang J. Investigating function of long noncoding RNA of HOTAIRM1 in progression of SKOV3 ovarian cancer cells. Drug Dev Res. 2021; 82: 1162-1168.
- Chao H, Zhang M, Hou H, Zhang Z, Li N. HOTAIRM1 suppresses cell proliferation and invasion in ovarian cancer through facilitating ARHGAP24 expression by sponging miR-106a-5p. Life Sci. 2020; 243: 117296.
- Zheng M, Liu X, Zhou Q, Liu G. HOTAIRM1 competed endogenously with miR-148a to regulate DLGAP1 in head and neck tumor cells. Cancer Med. 2018; 7: 3143-3156.
- Zhang Y, Mi L, Xuan Y, Gao C, Wang YH, Ming HX, Liu J. LncRNA HOTAIRM1 inhibits the progression of hepatocellular carcinoma by inhibiting the Wnt signaling pathway. Eur Rev Med Pharmacol Sci. 2018; 22: 4861-4868.
- Chen TJ, Gao F, Yang T, Li H, Li Y, Ren H, Chen MW. LncRNA HOTAIRM1 Inhibits the Proliferation and Invasion of Lung Adenocarcinoma Cells via the miR-498/WWOX Axis. Cancer Manag Res. 2020; 12: 4379-4390.
- Xu F, Chen M, Chen H, Wu N, Qi Q, Jiang X, Fang D, Feng Q, Jin R, Jiang L. The Curcumin Analog Da0324 Inhibits the Proliferation of Gastric Cancer Cells via HOTAIRM1/miR-29b-1-5p/PHLPP1 Axis. J Cancer. 2022; 13: 2644-2655.
- Lu R, Zhao G, Yang Y, Jiang Z, Cai J, Zhang Z, Hu H. Long noncoding RNA HOTAIRM1 inhibits cell progression by regulating miR-17-5p/ PTEN axis in gastric cancer. J Cell Biochem. 2019; 120: 4952-4965.
- Wan L, Kong J, Tang J, Wu Y, Xu E, Lai M, Zhang H. HOTAIRM1 as a potential biomarker for diagnosis of colorectal cancer functions the role in the tumour suppressor. J Cell Mol Med. 2016; 20: 2036-2044.
- Ren T, Hou J, Liu C, Shan F, Xiong X, Qin A, Chen J, Ren W. The long non-coding RNA HOTAIRM1 suppresses cell progression via sponging endogenous miR-17-5p/ B-cell translocation gene 3 (BTG3) axis in 5-fluorouracil resistant colorectal cancer cells. Biomed Pharmacother. 2019; 117: 109171.
- Salido-Guadarrama I, Romero-Cordoba SL, Rueda-Zarazua B. Multi-Omics Mining of lncRNAs with Biological and Clinical Relevance in Cancer. Int J Mol Sci. 2023; 24: 16600.
- Mahato RK, Bhattacharya S, Khullar N, Sidhu IS, Reddy PH, Bhatti GK, Bhatti JS. Targeting Long Non-Coding RNAs in Cancer Therapy using CRISPR-Cas9 Technology: A Novel Paradigm for Precision Oncology. J Biotechnol. 2023; 0168-1656: 00213-4.
- Yang Q, Fu Y, Wang J, Yang H, Zhang X. Roles of lncRNA in the diagnosis and prognosis of triple-negative breast cancer. J Zhejiang Univ Sci B. 2023; 24: 1123-1140.
- Alharthi NS, Al-Zahrani MH, Hazazi A, Alhuthali HM, Gharib AF, Alzahrani S, Altalhi W, Almalki WH, Khan FR. Exploring the lncRNA-VEGF axis: Implications for cancer detection and therapy. Pathol Res Pract. 2023; 253: 154998.
- Mehmandar-Oskuie A, Jahankhani K, Rostamlou A, Mardafkan N, Karamali N, Razavi ZS, Mardi A. Molecular mechanism of lncRNAs in pathogenesis and diagnosis of auto-immune diseases, with a special focus on lncRNA-based therapeutic approaches. Life Sci. 2023; 336: 122322.
- Li Q, Dong C, Cui J, Wang Y, Hong X. Over-expressed lncRNA HOTAIRM1 promotes tumor growth and invasion through up-regulating HOXA1 and sequestering G9a/EZH2/Dnmts away from the HOXA1 gene in glioblastoma multiforme. J Exp Clin Cancer Res. 2018; 37: 265.
- Kim CY, Oh JH, Lee JY, Kim MH. The LncRNA HOTAIRM1 Promotes Tamoxifen Resistance by Mediating HOXA1 Expression in ER+ Breast Cancer Cells. J Cancer. 2020; 11: 3416-3423.
- Wang XQD, Dostie J. Reciprocal regulation of chromatin state and architecture by HOTAIRM1 contributes to temporal collinear HOXA gene activation. Nucleic Acids Res. 2017; 45: 1091-1104.
- Usman M, Li A, Wu D, Qinyan Y, Yi LX, He G, Lu H. The functional role of lncRNAs as ceRNAs in both ovarian processes and associated diseases. Noncoding RNA Res. 2023; 9: 165-177.
- Kim S. LncRNA-miRNA-mRNA regulatory networks in skin aging and therapeutic potentials. Front Physiol. 2023; 14: 1303151.
- Lin Y, Wen H, Yang B, Wang C, Liang R. Integrated bioinformatics and validation to construct lncRNA-miRNA-mRNA ceRNA network in status epilepticus. Heliyon. 2023; 9: 22205.
- Jiang M, Wang Z, Lu T, Li X, Yang K, Zhao L, Zhang D, Li J, Wang L. Integrative analysis of long noncoding RNAs dysregulation and synapse-associated ceRNA regulatory axes in autism. Transl Psychiatry. 2023; 13: 375.
- Wang X, Liu Y, Lei P. LncRNA HOTAIRM1 promotes osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by targeting miR-152-3p/ETS1 axis. Mol Biol Rep. 2023; 50: 5597-5608.
- Wang G, Yu Y, Wang Y. Effects of propofol on neuroblastoma cells via the HOTAIRM1/miR-519a-3p axis. Transl Neurosci. 2022; 13: 57-69.
- Zhou Y, Wu Q, Long X, He Y, Huang J. lncRNA HOTAIRM1 Activated by HOXA4 Drives HUVEC Proliferation Through Direct Interaction with Protein Partner HSPA5. Inflammation. 2023.
- Han W, Wang S, Qi Y, Wu F, Tian N, Qiang B, Peng X. Targeting HOTAIRM1 ameliorates glioblastoma by disrupting mitochondrial oxidative phosphorylation and serine metabolism. iScience. 2022; 25: 104823.
- Chen W, Liu J, Ge F, Chen Z, Qu M, Nan K, Gu J, Jiang Y, Gao S, Liao Y. Long Noncoding RNA HOTAIRM1 Promotes Immunosuppression in Sepsis by Inducing T Cell Exhaustion. J Immunol. 2022; 208: 618-632.
- Herman AB, Tsitsipatis D, Gorospe M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol Cell. 2022; 82: 2252-2266.
- Peng WX, Koirala P, Mo YY. LncRNA-mediated regulation of cell signaling in cancer. Oncogene. 2017; 36: 5661-5667.
- Ge T, Gu X, Jia R, Ge S, Chai P, Zhuang A, Fan X. Crosstalk between metabolic reprogramming and epigenetics in cancer: updates on mechanisms and therapeutic opportunities. Cancer Commun (Lond). 2022; 42: 1049-1082.
- Guo H, Li T, Sun X. LncRNA HOTAIRM1, miR-433-5p and PIK3CD function as a ceRNA network to exacerbate the development of PCOS. J Ovarian Res. 2021; 14.
- Dahariya S, Raghuwanshi S, Thamodaran V, Velayudhan SR, Gutti RK. Role of Long Non-Coding RNAs in Human-Induced Pluripotent Stem Cells Derived Megakaryocytes: A p53, HOX Antisense Intergenic RNA Myeloid 1, and miR-125b Interaction Study. J Pharmacol Exp Ther. 2023; 384: 92-101.
- Li YR, Yan WJ, Cai LL, Deng XL. Effect of Down-Regulation of LncRNA-HOTAIRM1 to Proliferation, Apoptosis and KIT/AKT Pathway of Jurkat Cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2021; 29: 1123-1128.s
- Liu WB, Li GS, Shen P, Li YN, Zhang FJ. Long non-coding RNA HOTAIRM1-1 silencing in cartilage tissue induces osteoarthritis through microRNA-125b. Exp Ther Med. 2021; 22: 933.
- Tollis P, Vitiello E, Migliaccio F, D’Ambra E, Rocchegiani A, Garone MG, Bozzoni I, Rosa A, Carissimo A, Laneve P, et al. The long noncoding RNA nHOTAIRM1 is necessary for differentiation and activity of iPSC-derived spinal motor neurons. Cell Death Dis. 2023; 14: 741.
- Segal D, Coulombe S, Sim J, Josée Dostie J. A conserved HOTAIRM1-HOXA1 regulatory axis contributes early to neuronal differentiation. RNA Biol. 2023; 20: 1523-1539.
- Gu D, Tong M, Wang J, Zhang B, Liu J, Song G, Zhu B. Overexpression of the lncRNA HOTAIRM1 promotes lenvatinib resistance by downregulating miR-34a and activating autophagy in hepatocellular carcinoma. Discov Oncol. 2023; 14: 66.
- Liang L, Gu W, Li M, Gao R, Zhang X, Guo C, Mi S. The long noncoding RNA HOTAIRM1 controlled by AML1 enhances glucocorticoid resistance by activating RHOA/ROCK1 pathway through suppressing ARHGAP18. Cell Death Dis. 2021; 12: 702.
- Chen L, Hu N, Wang C, Zhao H. HOTAIRM1 knockdown enhances cytarabine-induced cytotoxicity by suppression of glycolysis through the Wnt/β-catenin/PFKP pathway in acute myeloid leukemia cells. Arch Biochem Biophys. 2020; 680: 108244.