
Journal of Clinical Images and Medical Case Reports
ISSN 2766-7820
Research Article - Open Access, Volume 3
Research status of vitiligo models
Yi-lu Zhao1,2; Ge Kang1,2; Ru-zhi Zhang2*
1Bengbu Medical University, Bengbu, Anhui, 233000, China.
2Department of Dermatology, The Third Affiliated Hospital of Soochow University, Changzhou, 213003, China.
*Corresponding Author : Ru-zhi Zhang
Department of Dermatology, The Third Affiliated Hospital of Soochow University, Changzhou, Jiangsu 213003, China.
Email: zhangruzhi628@163.com
Received : Jul 21, 2022
Accepted : Aug 09, 2022
Published : Aug 16, 2022
Archived : www.jcimcr.org
Copyright : ©Ru-zhi Z (2022).
Abstract
Vitiligo is a multifactorial autoimmune disease characterized by depigmentation of the skin and mucous membranes. The etiology and pathogenesis of vitiligo are not yet fully understood. The establishment of suitable vitiligo models will facilitate studies of the pathogenesis and therapeutic treatments of vitiligo. At present, vitiligo models have usually been established by treatment with chemical substances such as monobenzone and hydroquinone, and by the exogenous injection of CD8+ T cells into transgenic mice. Those models have their own advantages and disadvantages, and are suitable for different types of research. In addition to animal models, many drug-induced types of vitiligo have been found clinically, such as vitiligo-like skin lesions caused by some chemotherapy drugs, antiepileptic drugs and antifungal drugs. This article will describe some methods used to establish vitiligo models, to detect biomarkers associated with vitiligo, and will discuss the application of vitiligo models.
Keywords: Vitiligo; Animal; Model; Medicated-associated.
Citation: Zhao YL, Kang G, Ru-zhi Z. Research status of vitiligo models. J Clin Images Med Case Rep. 2022; 3(8): 2001.
Background
Vitiligo is an autoimmune skin disease with an approximate prevalence of 0.5-2% worldwide that is characterized by chalky-white macules and/or patches with distinct margins [1]. Depigmented spots of the skin can appear anywhere on the body. Although vitiligo does not affect the lifespan of the patient or the function of important organs, the impact on the patient’s quality of life is obvious, even devastating. Research on the pathogenesis and treatment of vitiligo has never stopped or slowed down. The establishment of animal models for various diseases is an important means of modern medical research. Establishing an animal model of vitiligo that is highly similar to the clinical manifestations of patients is crucial for the development of effective therapeutic drugs. This article reviews recent studies of vitiligo models in order to promote in-depth research on vitiligo.
Chemical vitiligo
Initially, chemical-induced skin depigmentation was considered an occupational hazard. Later, it was found that some commonly used daily necessities, such as disinfectants, diaper creams, cosmetic dyes, etc., can also cause skin discolorations. These chemically-induced depigmented spots were named “chemical vitiligo” by Ghosh, which is defined as acquired depigmentation caused by repeated exposure to a specific compound in genetically susceptible individuals, which is similar to idiopathic vitiligo [2].
The main mechanisms underlying chemical vitiligo include [3]: (1) The common structure of offending chemicals in the majority of this category is a phenol group, which is shared by tyrosine, the amino acid that is the basis of melanin synthesis. Depigmenting chemicals appear to disrupt melanin production by interacting with tyrosinase (TYR), which leads to the development of depigmented spots. (2) Depigmenting chemicals can act to induce the death of melanocytes (MCs) indirectly by activating an inflammatory cascade in dendritic cells. (3) The chemical-induced stress initiates autophagy, which induces the production of exosomes and leads to an autoimmune response that results in the destruction of MCs (Figure 1). Substances that have been reported to cause chemical vitiligo directly or indirectly include: Monobenzone (MBZ), P-Phenylenediamine (PPD) and Hydrogen Peroxide (H2O2). Those chemicals have been used to treat severely generalized vitiligo patients to remove remnants of normal skin tone and improve the patient’s overall appearance.
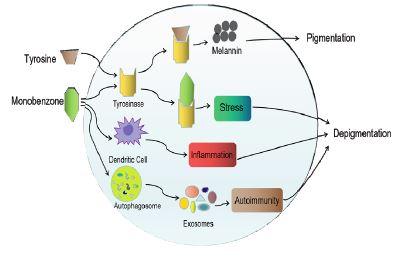
Monobenzone (MBZ)
In 1939, Oliver et al. found that some workers in a leather manufacturing company had discolored spots on their hands and forearms covered by their gloves and on remote places of their bodies not covered by their gloves [4]. To further explore whether MBZ can induce depigmentation of non-exposed skin sites in mice, Zhu et al [5]. Used 4-week-old male C57BL/6 mice and treated them topically with MBZ cream. They found that small white patches first appeared in the drug contact area and gradually expanded. After prolonged application of MBZ, some mice also developed depigmented spots on untreated areas such as their trunk, ears and/or tail. Over time, the areas of depigmented spots expanded and eventually lead to depigmentation of most parts of the body. The proportion of mice that developed distant depigmentation and the severity of the depigmentation increased with higher MBZ concentrations. Ultimately, all MBZ-treated mice developed depigmentation in exposed areas and in some non-exposed areas. High concentrations of MBZ can shorten the time until the appearance of vitiligo, but there is also significant local inflammation and exudation. Therefore, they concluded that a 40% MBZ cream was the most effective concentration. Reflectance Confocal Microscopy (RCM) and histological examination of the depigmented skin showed the loss of epidermal MCs and the accumulation of CD8+ T cells around the lesions, with elevated skin mononuclear cell infiltrations around the distant depigmented lesions. Immunofluorescence staining with an anti-CD8 antibody showed that the infiltrating lymphocytes included a large number of CD8+ T cells, while only a small number of CD8+ T cells were observed in healthy skin [6]. These results suggested that the appearance of distal leukoplakia in mice is related to the immune response of T cells.
Hydrogen peroxide (H2O2)
H2O2 is a two-electron reduction product with strong oxidizing properties and is one of the raw materials for generating hydroxyl radicals. H2O2 can pass through the cell membrane and can directly cause damage to MCs. H2O2 can also affect melanin production by inhibiting the Tyrosine Hydroxylase (TH) and Dopa Oxidase (DO) activities of tyrosinase and by oxidizing tetrahydrobiopterin (BH-4) in the epidermis. After exposure to H2O2, MCs had decreased Bcl-2 levels and increased levels of Bax and cleaved Caspase-3. Various endogenous or exogenous stimuli can disrupt the balance of the oxidative-antioxidant system, which may increase the level of Reactive Oxygen Species (ROS) in MCs. ROS can destroy macromolecular substances including lipids, nucleic acids and proteins, leading to lipid peroxidation, DNA fragmentation, oxidative decomposition of proteins and activation or inactivation of various enzymes [7]. Moreover, ROS may also cause a decrease in mitochondrial membrane potential, abnormalities in mitochondrial membrane lipid composition, and impairment of the integrity of the respiratory chain. Those conditions make MCs more sensitive to apoptosis stimuli and more easily damaged. Furthermore, oxidative stress disrupts the folding machinery of the endoplasmic reticulum and activates the Unfolded Protein Response (UPR). If the adaptive mechanisms in MCs cannot resolve the protein-folding defect, the cells undergo apoptosis. The amount of E-cadherin in the cell membrane is down-regulated due to oxidative damage, resulting in impaired adhesion of MCs to the basement membrane and triggering the apoptosis of MCs [8].
One research study applied a 5% H2O2 solution at a depilated area of about 40 cm2 on the dorsal skin of black guinea pigs, 0.5 ml each time, twice a day for 50 days [9]. During that time, the black skin of the guinea pigs turned white and even grew white hair. Local histopathological examination showed that melanin content had decreased in the spinous layer and in Hair Follicles (HFs). Some HFs were white in the upper part, yellow in the middle, and black in the lower part. At the same time, serum levels of Monoamine Oxidase (MAO) and Immunoglobulin (Ig) were increased, and Cholinesterase (ChE) decreased. However, a few research studies failed to induce vitiligo in mice using H2O2. They speculated that H2O2 achieves depigmentation through the bleaching of existing melanin. The effect of H2O2 on MCs can be offset by more newly generated melanin granules. The H2O2-induced depigmentation is easy to operate, but it is not completely consistent with the pathogenesis of human vitiligo.
Hydroquinone (HQ)
HQ is frequently used as a skin lightening agent that acts by inhibiting the TYR-catalyzed conversion of tyrosine to melanin [10]. The mechanism of toxicity of HQ on MCs is unclear. Possible mechanisms include [11]: (1) rapid spontaneous oxidation of HQ in the extracellular environment to generate the corresponding o-quinones and abundant ROS, (2) hydrophobic electron-donating substitutes at the para-position of phenol, conferring high cellular permeability and high sensitivity to auto-oxidation. Studies have shown that the clinical use of a 2% HQ cream can effectively reduce melanin production by 63.5%. Topical treatment with 2 mL 2.5% HQ was applied to shaved areas (2 × 2 cm) of the dorsal skin of C57BL/6 mice daily for 60 days. Eventually, white spots and hair appearedon the treated areas. Histological analysis showed that the amount of melanin in MCs in HFs, in basal MCs and in epidermal MCs were all reduced after HQ treatment compared to the control group.
Immune-related vitiligo animal models
Smyth Line (SL) chickens
Smyth Line (SL) chickens are the only animal model of vitiligo that spontaneously display all clinical and biological manifestations of human vitiligo. SL vitiligo involves the interaction of genetic, immune and environmental factors. Susceptibility to SL vitiligo is manifested in part in an inherent defect of MCs. The loss of MCs is due to MC-specific cell-mediated immune responses and abnormalities in humoral immunity. The phenotype and etiology of SL vitiligo is similar to human vitiligo [12]. However, the loss of pigmentation in SL chickens is due to the loss of MCs in feathers and pigmented HFs without involving the loss of epidermal pigmentation [13]. Therefore, SL chickens are not suitable for studies of human vitiligo.
Mouse models of vitiligo
Studies have shown that T lymphocytes, mainly cytotoxic CD8+ T cells, that infiltrate the edges of vitiligo lesions, mediate the elimination of MCs and play a necessary and sufficient role in the occurrence and development of vitiligo. While regulatory T cells (Tregs) maintain immune homeostasis and suppress inflammation, skin Resident Memory T (TRM) cells induce vitiligo relapse [14]. According to this theory, several kinds of mouse vitiligo models have been established.
Transgenic mice: Transgenic mice (KRT14-kitl*4XTG2Bjl, named SCF mice) that express Stem Cell Factor (SCF) induced by the Kit ligand are used as recipient mice, whose hairs and epidermis are black. Those mice were irradiated with at a sublethal dose (5 Gy) of gamma rays to deplete endogenous lymphocytes. DPEGEP mice which express green fluorescent protein (GFP) were crossed with T Cell Receptor (TCR) transgenic mice expressing Premelanosome Protein (PMEL) to obtain GFP-PMEL TCR transgenic mice (referred to as PMEL mice), who served as a donor of CD8+ T cells. After the intraorbital injection of CD8+ T cells into SCF mice, an attenuated recombinant Vaccinia Virus (rVV) was intraperitoneally injected to activate CD8+ T cells to specifically kill MCs of the recipient mice. Five to seven weeks later, the SCF mice developed depigmented skin, mostly on their ears, nose, palms, soles and tails. No additional depigmented spots appeared after 7 weeks [16]. This model of transgenic mice is similar to the progression of human vitiligo, preserving the important role of HFs in repigmentation, and therefore it is suitable for research studies on the mechanism of repigmentation and on the efficacy of new drugs for treating vitiligo.
C57BL/6 mice: According to the phenomenon whereby MCs are killed by the body’s immune cells during the treatment of Malignant Melanoma (MM), research groups have constructed a rVV expressing a variety of MC-specific antigens [17]. When the rVV expressing human TRP-1 was injected into C57BL/6 mice, 80% of them developed depigmented skin patches, which were similar to the vitiligo-like depigmented patches seen in human MM patients treated with IL-12 chemotherapy [18]. The establishment of this model is due to the activation of CD4+ T cell-mediated cellular immunity and the production of specific antibodies that result in the damage of MCs. This mouse model of vitiligo can be used to study the immune-related mechanisms of vitiligo.
Hybrid mice: By hybridization of TYR-deficient albino mice with AAD+ mice expressing type I Major Histocompatibility Complex (MHC), human HLA-A*021 binding site and TYR369 phenotype, the model of TYR369-specific transgenic mice (called FH mice) was constructed [19]. The FH mice were then crossed with AAD+ mice, and depigmentation of the hair and skin appeared in their offspring. This model is a CD8+ T cell-dependent autoimmune vitiligo, which is similar to human vitiligo. In this model, the depigmented spots appeared slowly, with bilateral lesions on the cephalic and facial regions in young animals and on the trunk area only in adult animals. This model suggests that the process of depigmentation may be mechanistically different in different regions of the body. This model could be used to further characterize the immune mechanisms of vitiligo involving cytokines, immune pathways and regional mechanisms.
Medicinal-associated vitiligo
Depigmented patches associated with melanoma have various phenotypes, including white halos around the primary tumor, white areas mixed with the melanoma, and nonpigmented patches at melanoma scars or at the sites of immunotherapy injections [20]. The rarest manifestations are vitiligo-like patches distant from the primary tumor. Immunotherapy probably increases the incidence of vitiligo associated with melanoma. Because MM cells and MCs share a variety of immunogenic antigens, including MELAN-A/Mart-1, TYRP-1/GP75, TYRP-2, TYR and GP100, the autoimmune-mediated destruction of MM cells can also lead to the killing of normal MCs, resulting in the depigmentation [21]. Research studies have found that spontaneous vitiligo is more common in MM patients than in the general population. Moreover, following treatment with immunosuppressants such as Interferon Α (IFN-α), IL-2 or immune checkpoint inhibitors (anti-cytotoxic T-lymphocyte antigen-4 and programmed cell death inhibitors), 2 to 16% of MM patients develop depigmented spots, which is referred as MM-Associated Vitiligo (MAV), which shares similar pathological features to vitiligo. The phenomenon of MAV may predict a better immune response, which means better survival than expected according to the melanoma stage [22].
Vitiligo-like depigmentation can also occur during or after the treatment of other diseases. Imiquimod cream is routinely used to treat several benign and malignant skin diseases due to its antiviral, antitumor and immunomodulatory properties, and imiquimod induced local hypopigmentation in some patients by inhibiting melanogenesis, activating autophagy, increasing the apoptosis of MCs and stimulating the secretion of proinflammatory cytokines including interleukin 6 (IL‐6), IL‐8 and soluble ICAM‐1 (sICAM‐1) [23]. It has been reported that a patient with cardiovascular disease developed depigmented spots on the back of the hand, the wrist, waist and abdomen after 6 days of taking Xuesaitong capsule which which is a Chinese herbal monomer preparation consisting of Panax notoginseng saponins [24]. Furthermore, Patients with breast cancer developed vitiligo-like depigmentation (DI-VLD) associated with treatment by an inhibitor of cyclin-dependent kinases 4 and 6 (CDK 4/6) [25].
Conclusion
Currently, the animals used to establish vitiligo models include Smyth chickens, mice, Sinclair pigs, dogs, cats, gray horses, rats and zebrafish. Different animal models have their own advantages and limitations, and can only simulate the pathogenesis of human vitiligo to a certain extent. Researchers should choose different models to conduct experiments according to their own research directions. In the future, researchers should also be committed to developing better vitiligo models closer to the human body environment, so as to better understand the pathogenesis of vitiligo and develop efficient and safe treatment methods for vitiligo.
Declarations
Authors’ contributions: Author Yi-luzhao prepared and wrote the manuscript. Author Ge Kang provided imaging and edited the manuscript. Author Ru-zhi Zhang revised it critically. All authors read and approved the final version of the manuscript.
Acknowledgments: The authors are very grateful to Professor V.J. Hearing for help with the English language editing.
Funding: The work was supported by the Foundation of the National Natural Science (No: 81673078) and the Graduate Research Innovation Program of Bengbu Medical College (Byycx21088).
References
- Zhang J, Hu W, Wang P, Ding Y, Wang H , et al. Research Progress on Targeted Antioxidant Therapy and Vitiligo. Oxid Med Cell Longev. 2022; 2022: 1821780.
- Ghosh S. Chemical Vitiligo: A Subset of Vitiligo. Indian J Dermatol. 2020; 65: 443-449.
- Gupta D, Kumari R and Thappa DM. Depigmentation therapies in vitiligo. Indian J Dermatol Venereol Leprol. 2012; 78: 49-58.
- Kaushik H, Kaul D, Kumaran MS and Parsad D. Chemical induced pathognomonic features observed in human vitiligo are mediated through miR-2909 RNomics pathway. J Dermatol Sci. 2020; 100: 92-98.
- Zhu Y, Wang S and Xu A. A mouse model of vitiligo induced by monobenzone. Exp Dermatol. 2013; 22: 499-501.
- Arowojolu OA, Orlow SJ, Elbuluk N and Manga P. The nuclear factor (erythroid-derived 2)-like 2 (NRF2) antioxidant response promotes melanocyte viability and reduces toxicity of the vitiligo-inducing phenol monobenzone. Exp Dermatol. 2017; 26: 637-644.
- An R, Li D, Dong Y, She Q, Zhou T, et al. Methylcobalamin Protects Melanocytes from H2O2-Induced Oxidative Stress by Activating the Nrf2/HO-1 Pathway. Drug Des DevelTher. 2021; 15: 4837-4848.
- Xuan Y, Yang Y, Xiang L and Zhang C. The Role of Oxidative Stress in the Pathogenesis of Vitiligo: A Culprit for Melanocyte Death. Oxid Med Cell Longev. 2022; 2022: 8498472.
- Sun MC, Xu XL, Du Y, Lou XF, Wang W, et al. Biomimetic Melanosomes Promote Orientation-Selective Delivery and Melanocyte Pigmentation in the H2O2-Induced Vitiligo Mouse Model. ACS Nano. 2021.
- Hu ZM, Zhou Q, Lei TC, Ding SF, Xu SZ, et al. Effects of hydroquinone and its glucoside derivatives on melanogenesis and antioxidation: Biosafety as skin whitening agents. J Dermatol Sci. 2009; 55: 179-184.
- Huo SX, Liu XM, Ge CH, Gao L, Peng XM, et al. The effects of galangin on a mouse model of vitiligo induced by hydroquinone. Phytother Res. 2014; 28: 1533-1538.
- Sreekumar GP, Smyth JR Jr and Ponce de Leon FA. Molecular characterization of the Smyth chicken sublines and their parental controls by RFLP and DNA fingerprint analysis. Poult Sci. 2001; 80: 1-5.
- Erf GF, Bersi TK, Wang X, Sreekumar GP and Smyth JR Jr. Herpesvirus connection in the expression of autoimmune vitiligo in Smyth line chickens. Pigment Cell Res. 2001; 14: 40-46.
- Erf GF, Le Pape G, Rémy S and Denesvre C. Mardivirus Infection and Persistence in Feathers of a Chicken Model Harboring a Local Autoimmune Response. Microorganisms. 2020; 8: 1613.
- Wang Y, Li S and Li C. Clinical Features, Immunopathogenesis, and Therapeutic Strategies in Vitiligo. Clin Rev Allergy Immunol. 2021; 61: 299-323.
- Riding RL, Richmond JM and Harris JE. Mouse Model for Human Vitiligo. CurrProtoc Immunol. 2019; 124: e63.
- catozza F, Moschella F, D’Arcangelo D, Rossi S, Tabolacci C, et al. Nicotinamide inhibits melanoma in vitro and in vivo. J Exp Clin Cancer Res. 2020; 39: 211.
- Steitz J, Tüting T. Biolistic DNA vaccination against melanoma. Methods Mol Biol. 2013; 940: 317-337.
- Gregg RK, Nichols L, Chen Y, Lu B, Engelhard VH, et al. Mechanisms of spatial and temporal development of autoimmune vitiligo in tyrosinase-specific TCR transgenic mice. J Immunol. 2010; 184: 1909-1917.
- Daneshpazhooh M, Shokoohi A, Dadban A, Raafat J. The course of melanoma-associated vitiligo: report of a case. Melanoma Res. 2006; 16: 371-373.
- Teulings HE, Willemsen KJ, Glykofridis I, Krebbers G, Komen L, et al. The antibody response against MART-1 differs in patients with melanoma-associated leucoderma and vitiligo. Pigment Cell Melanoma Res. 2014; 27: 1086-1096.
- Farinazzo E, Zelin E, Agozzino M, Papa G, Pizzichetta MA, et al. Regression of nevi, vitiligo-like depigmentation and halo phenomenon may indicate response to immunotherapy and targeted therapy in melanoma. Melanoma Res. 2021; 31: 582-585.
- Piqué Duran E. Recurrence of a Basal Cell Carcinoma in a Vitiligo-like Area after Imiquimod Treatment. ActasDermosifiliogr. 2021; 112 Suppl 1: 14. English, Spanish.
- Chen J, Cen X and Shi J. Vitiligo-like depigmentation associated with Xuesaitong capsule treatment. Australas J Dermatol. 2021; 62: e338-e339.
- Sollena P, Nikolaou V, Soupos N, Kotteas E, Voudouri D, et al. Vitiligo-like lesions in patients with advanced breast cancer treated with cycline-dependent kinases 4 and 6 inhibitors. Breast Cancer Res Treat. 2021; 185: 247-253.