
Journal of Clinical Images and Medical Case Reports
ISSN 2766-7820
Research Article - Open Access, Volume 4
OX-LDL promotes insufficient autophagy and apoptosis of
ovarian granulosa cells through regulation ROS-mediated
PI3K/Akt/mTOR pathway
Fengping He*; Yanhui Liu; Zhang Yongmei
Affiliated Dongguan Maternal and Child Health Hospital, Southern Medical University, Guangzhou, China.
*Corresponding Author : Fengping He
Affiliated Dongguan Maternal and Child Health Hospital, Southern Medical University, Guangzhou, China.
Email: 1458574250@qq.com;
fengphe@hotmail.com
Received : Feb 28, 2023
Accepted : Mar 16, 2023
Published : Mar 23, 2023
Archived : www.jcimcr.org
Copyright : © He F (2023).
Abstract
The purpose of this study was to determine the role of Oxidized Low-Density Lipoprotein (OX-LDL) promotes insufficient autophagy and apoptosis of ovarian granulosa cells through regulation Reactive Oxygen Species (ROS)-mediated PI3K/Akt/mTOR pathway. In vivo studies, we established mouse models of OX-LDL induced ovarian dysfunction and tested the changes of follicle-stimulating hormone (FSH) and estradiol (E2) under OX-LDL. In the current study, it showed significantly reduced autophagy level and increased apoptosis of ovarian granulosa cells in OX-LDL- treated C57BL/6 mice. Moreover, western blotting showed that OX-LDL treatment markedly down-regulation phosphoinositide 3- kinase (PI3K), threonine- protein kinase (Akt) and mammalian target of rapamycin (mTOR) expression by ROS-mediated. Additionally, OX-LDL treatment significantly promoted the gene and protein expression of caspase 3. In the present study, we found that 100-400 μg/mL OX-LDL dose-dependently enhanced ROS levels, induced granulocellular apoptosis, and down-regulated the expression of autophagic markers Beclin1, and the ratio of LC3-II:LC3-I decreased. Western blotting demonstrated that OX-LDL could not only regulate the ROS-mediated PI3K/Akt/mTOR signaling pathway, but also stimulate the expression of apoptosis- related gene (caspase 3). In addition, we proved that miRNA-146 pretreatment could inhibit OX-LDL regulated ROS-mediated PI3K/Akt /mTOR signaling pathway. miRNA-146 also suppressed the expression of caspase 3 under OX-LDL treatment. In conclusion, OX-LDL can promote insufficient autophagy and apoptosis of ovarian granulosa cells by regulating ROS-mediated PI3K/Akt/mTOR pathway in Primary ovarian insufficiency (POI) mouse models.
Keywords: Primary ovarian insufficiency; Insufficient autophagy; Oxidized low density lipoprotein; Reactive oxygen species; Phosphoinositide 3- kinase (PI3K)/ serine/threonine-protein kinase (Akt)/ mammalian target of rapamycin (mTOR).
Citation: He F, Liu Y, Yongmei Z. OX-LDL promotes insufficient autophagy and apoptosis of ovarian granulosa cells through regulation ROS-mediated PI3K/Akt/mTOR pathway. J Clin Images Med Case Rep. 2023; 4(3): 2337.
Introduction
Primary Ovarian Insufficiency (POI) usually refers to amenorrhea in women before 40 years of age, with elevated serum follicle stimulating hormone (FSH) and decreased estrogen (E2) levels [1,2]. Loss of ovarian function leads to amenorrhea and atrophy of sexual organs, and inhibits follicles, growth and development [2]. POI also includes premature ovarian failure (POF), which is the final stage of POI development [3]. The etiology of POI is still unknown, and its pathogenesis and pathogenesis are very complex, which has not been clarified at present. Recent studies have shown that oxidative stress is also related to human reproductive system diseases [4]. In POI, inflammation reaction and oxidative stress affect the quality of follicles [5]. Oxidative stress during ovarian aging is an important pathogenesis [6]. Oxidized Low-Density Lipoprotein (OX-LDL) is thought to be a key factor in inducing oxidative stress. Oxidative stress and OX-LDL may affect redox status and/or redox sensitive signaling pathways and gene expression, change proteins and DNA, and promote apoptosis [7]. It was also found that embryonic, follicular, and ovarian granulosa cells increased Reactive Oxygen Species (ROS) expression and oxidative stress induced OX-LDL. Low levels of ROS and/or mild oxidative stress are involved in physiological processes, including cell signaling systems, cell growth and differentiation [8]. In addition, ROS levels in specific tissues lead to different processes of cell death. For example, increased ROS levels leads to granulosa cell apoptosis [9].
In humans, granulosa cells exposure to OX-LDL causes autophagy, which further leads to apoptosis and death. According to the study, POI has excess OX-LDL resulting in enhanced granulosa autophagy dysregulation and cell apoptosis or cell death [10]. However, the underlying mechanism through which autophagy dysregulation is involved in the apoptosis or cell death of granulosa cells remains to be determined. In this study, we demonstrated that under OX-LDL exposure ovarian granulosa cells insufficient autophagy and apoptosis coexist in the POI mouse models. Our findings provide the first evidence for the role of OX-LDL causes damage and deficiency of autophagy in granulosa cells and suggest that impaired autophagy in granulosa cells may be involved in the pathogenesis of POI. Furthermore, the regulation of ROS-mediated PI3K/Akt/mTOR pathway might be involved in OX-LDL induced autophagy dysregulation and apoptosis of ovarian granulosa cells.
Materials and methods
Animals
Female BALB/c mice (8 weeks; 18–22 g) were obtained from the Animal Laboratory of Sun Yat sen University. The mice were fed in an environment with temperature of 22 ± 2°C, humidity of 50 ± 5%, light of 12 hours and darkness of 12 hours, and adaptive feeding for one week. All animal experiments were approved by the Animal Experiment Ethics Committee of Sun Yat Sen University (NO.SYSU- IACUC- 2020-B1236).
Establishment of mouse primary ovarian insufficiency models
According to references [11,12], Katharina et al., (2013) [11] and Ying Liu et al., (2019) [12], POI mouse model was established by injecting OX-LDL (150 mg/kg), (n = 20). The establishment of the model was carried out by the Animal Laboratory of Sun Yat sen University. The mice were through the caudal vena cava injection with OX-LDL (150 mg/kg) for 7 days. After 20 days, blood samples were collected in dry test tubes without coagulant to obtain the serum. The concentration of estradiol (E2) and FSH in blood samples was tested by Chemiluminescence.
Isolation of mice granulosa cells and OX-LDL treatment
According to references [11,12], mice ovarian granulosa cells were obtained from BALB/c mice (n = 6) of the Animal Laboratory of Sun Yat sen University (Guangzhou, China). The female BALB/c mice ovaries were quickly dissected under sterile conditions and put into pre-cooled PBS to remove the surrounding tissues and surface capsule. Under the anatomical microscope, the follicles were punctured with a syringe needle to release the mice ovarian granulosa cells into DMEM-F12 medium, which were blown and dispersed into a single suspended cell in a centrifugal tube. Second, the ovarian granulosacells were incubated at 37°C, 5% CO2 for 24 h, after which 1 mg/ml 0.25% trypsin and 0.02% EDTA were added. Subsequently, the ovarian granulosa cells were incubated in an 37°C, 5% CO2 for another 60 min and filter them with a 200 mesh stainless steel cell sieve. The cells were cultured inDulbecco’s modified Eagle’s medium (DMEM; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented with 10% fetal bovine serum (FBS; Sigma- Aldrich; Merck KgaA), 100 U/ml penicillin G (Shanghai, China), 100 μg/ml streptomycin (Shanghai, China) and 2 mM glutamine (Guangzhou, China). OX-LDL was purchased from Shanghai (Shanghai Bioengineering Co., Ltd, China). Ovarian granulosa cells were cultured in CO2 incubator (37°C, saturation humidity, 5% CO2). Under serum-free conditions for 24, the granulosa cells were treated with OX-LDL under different conditions (100, 200, 300, 400 μg/mL).
MTT assay
The cell viability was analyzed by 3 – (4,5-dimethylthiazole-2-yl) – 2,5- diphenyl- 2-tetrazole ammonium bromide (MTT) colorimetry. In short, miR-146 transfected granulosa cells were inoculated at a density of 1 × 103 cells/well, and then the cells were cultured with OX-LDL for 24 hours. After rinsing twice with phosphate buffered saline (PBS), 10 μl MTT solution was added to each well with a final concentration of 5 mg/mL. The culture dish was cultured at 37°C for 10 min and 4 h, then 150 μl of dimethyl sulfoxide was added, and the absorbance was determined after shaking the plate for 10 min. Each experiment was repeated three times.
Apoptosis assay
The apoptosis of ovarian granulosa cells was analyzed using terminal deoxynuc- leotidyl transferase dUTP nick-end labeling (TUNEL) assay (Roche, Germany). For each sample, eight visual fields were randomly selected. The apoptotic index was calculated for 100 ovarian granulosa cells by dividing the number of apoptotic ovarian granulosa cells by the total number of ovarian granulosa cells.
ROS assay
The expression level of ROS in ovarian granulosa cells was detected by using probed with 10 μM Dichlorodihydrofluorescein diacetate (DCFH-DA, Beyotime Biotechnology, Shanghai, China) dissolved in serum-free medium at 37oC for 30 min after being treated with different concentrations of OX-LDL. The fluorescence was measured at 488 nm excitation/525 nm emission by the laser confocal scanning microscope (LCSM, Leica, Wetzlar, Germany). ROS in follicular fluid was detected by enzyme-linked immunosorbent assay (Shanghai Enzyme-linked Biotechnology Co., Ltd).
mTOR, PI3K, Akt and caspase 3 protein expression in ovarian granulosa cells were detected by western blot
Ovarian granulosa cells from ovarian tissues were treated by sonication, the lysates of cells were centrifuged and the proteins were separated by SDS-PAGE and then transferred to Immobilon NC membranes (Millipore, USA). After 2 h 5% skim milk blockage with Tris- buffered saline at room temperature, the membrane was incubated with primary antibodies against mTOR, PI3K, Akt and caspase 3, and β-actin (Abcam, UK) overnight at 4°C. Then, membranes were incubated with secondary antibodies conjugated with horseradish peroxidase for 1 h at 37°C. Blots were imaged using a Bio-Rad imaging system (Bio-Rad, USA).
Statistics
SPSS 19.0 was used to analyze the data which are presented as the mean ± standard error of the mean (SEM). One-way analysis of variance with a Bonferroni’s post hoc test was performed for multiple comparisons. P< 0.05 was considered to indicate a statistically significant difference.
Results
The expression of E2, FSH and ROS in POI model
Firstly, in this experiment, the concentration of estradiol (E2) and folicular stimulating hormone (FSH) was determined by using Chemiluminescence analyzed, respectively. The concentration of E2 was significantly reduced and FSH significantly enhanced in POI model (Table 1). Then, the levels expression of ROS in follicular fluid were measured by ELISA assay and significantly increased in POI model (Table 1).
Table 1: The course of endoscopic treatments.
| n | E2 (pmol/L) | FSH (U/L) | ROS (ng/ml) | |
|---|---|---|---|---|
| Control | 10 | 139.69 ± 25.87 | 12.76 ± 2.63 | 12.92 ± 4.72 |
| POI | 10 | 63.14 ± 17.18 | 61.43 ± 19.22 | 56.67 ± 23.84 |
| t | - | 19.286 | 17.795 | 13.669 |
| P | - | 0 | 0 | 0 |
Effects of OX-LDL on the viability of ovarian granulosa cells
To determine the appropriate dosage of OX-LDL used in this study, MTT assay was conducted. Cell viability of granulosa cells treated with OX-LDL at different concentrations (100, 200, 300 and 400 μg/mL) and at different times (12 h, 24 h, 48 h and 96 h). The number of ovarian granulosa cells decreased significantly at 96 h with 400 μg/mL OX-LDL treatment (P< 0.001) (Figure 1A). Therefore, the maximum concentration of OX-LDL used in the subsequent analysis was 400 μg/mL OX-LDL.
OX-LDL increased expression ROS levels during granulosa cells
Our previous research validated that OX-LDL has been proven to be associated with oxidative stress and over expression of ROS during inflammatory state. To investigate whether ROS was involved in OX-LD promoted granulosa cells insufficient autophagy and apoptosis increased, we determined the ROS levels in granulosa cells under OX-LDL. The results showed that OX-LDL-treatment led to increased fluorescence intensity of ROS in granulosa cells in a dose-dependent manner (Figure 1B). However, ROS generation decreased when using miRNA-146 mimics to neutralize intracellular ROS (Figure 1B). These findings suggested that intracellular ROS was activated in OX-LDL-promoted granulosa cells apoptosis (Figure 1F).
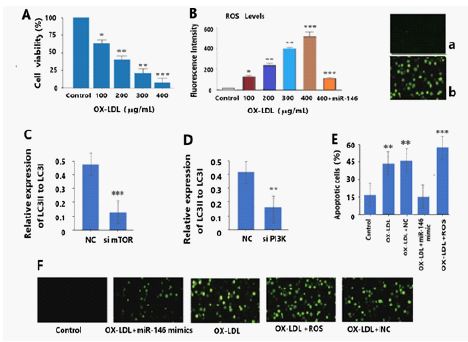
(A)The cell viability of granulosa cells after treatment with different concentrations (100, 200,300, and 400 μg/mL) of OX-LDL.
(B) OX-LDL treatment led to increased fluorescence intensity of ROS in granulosa cells in a dose-dependent manner, and miRNA-146 treatment decreased OX-LDL induced ROS generation (n = 5 per group), ***P< 0.001.
(C) The ratio of LC3II/LC3I was decreased in the si mTOR (0.13 ± 0.39) groups compared to the NC group (0.49 ± 0.56), Data are presented as mean ± SD, n = 5, ***P < 0.001 vs. NC.
(D) The ratio of LC3II/LC3I was decreased in the si PI3K (0.17 ± 0.46) groups compared to the NC group (0.42 ± 0.58), Data are presented as mean ± SD, n = 5, **P < 0.01 vs. NC.
(E) The apoptotic cells rate was measured by TUNEL assay. Percentage of apoptotic cells significantly decreased by expression of miRNA-146 compared with the OX-LDL group and OX-LDL+NC group, ** P < 0.01; *** P < 0.001.
(F) Up-regulation of miRNA-146 significantly reduced the apoptosis of OX-LDL treatment granulosa cells, while ROS remarkable enhanced the apoptosis of OX-LDL treatment granulosa cells. (a and b) : ROS level detected by DCFH-DA assay. OX-LDLsignificantly increased the ROS level. a, Control; b, OX-LDL
Insufficient autophagy inhibits granulosa cells activity
To determine the role of autophagy in granulosa cells, we used siRNA to knock down the expression of two key genes, mTOR, and PI3K, in granulosa cells of mouse (Figure 1C and 1D). The ratio of LC3-II to LC3-I expression was clearly decreased in both the si mTOR, and si PI3K, groups compared with the negative control (NC) group (Figure 1C and 1D), indicating the efficient inhibition of autophagy. Moreover, the accumulation of ROS also suggested the dysregulation of autophagy and apoptosis of granulosa cells (Figure 1E, F).
Effect of OX-LDL on autophagy markers and PI3K/Akt/mTOR in granulosa cells
As illustrated in Figure 2, the protein expressions of caspase 3 were dramatically up-regulated after OX-LDL stimulation in granulosa cells (Figure 2 A and D), while the protein expressions of p-PI3K, p-Akt, p-mTOR were dramatically down-regulated after OX-LDL stimulation in granulosa cells (Figure 3). Meanwhile, it could effectively decrease the ratio of LC3II/LC3I and Beclin1 with OX-LDL stimulation in granulosa cells (Figure 2 A, B and C). To confirm the involvement of ROS in the execution of PI3K/Akt/ mTOR pathway in OX-LDL-induced granulosa cells, we further investigated the protein expressions of the above molecules. Notably, miRNA-146 effectively promoted the up-regulations of the phosphorylated PI3K, Akt and mTOR caused by OX-LDL in vi tro (Figure 3A, B ,C and D). Interestingly, after ROS + miRNA-146 inhibitors co-incubation, the alterations were remarkably abrogated. These experimental data indicated the critical role of ROS in the regulation of PI3K/Akt/mTOR signalling Pathway.
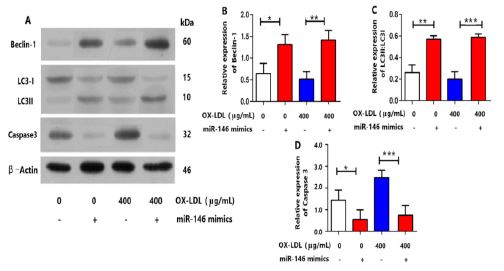
A. The protein expression levels of autophagy markers: LC3II/LC3I and Beclin1 were decreased under 400 μg/mL OX-LDL and promoted by miRNA-146 with 100 nmol/L. OX-LDL or without treatment as examined by Western blot (n = 5 per group). The protein expression levels of caspase 3 were increased under 400 μg/mL OX-LDL and restrained by miRNA-146 with 100 nmol/L. OX-LDL or without treatment as examined by Western blot (n = 5 per group).
B. Quantification of Beclin-1 in (A), * P< 0.05, ** P< 0.01.C. Quantification of LC3-I/LC3-II
in (A), ** P< 0.01,***P< 0.001.D. Quantification of caspase3 in (A), * P< 0.05, ***P< 0.001
Inhibition of ROS by miRNA-146 suppressed OX-LDL-induced PI3K/Akt/ mTOR expression
So far, we have proven that OX-LDL could induce insufficient autophagy and apoptosis of granulosa cells by regulating the mTOR expression. We also found that OX-LDL could increase ROS level in granulosa cells. Given that OX-LDL and ROS both regulated the mTOR expression and induced insufficient autophagy and apoptosis of granulosa cells increased and affect the quality of follicles, ROS might play a crucial role in linking OX-LDL and the PI3K/Akt/mTOR signaling pathway in insufficient autophagy and apoptosis of granulosa cells. Next, to determine whether OX-LDL-promoted insufficient autophagy and apoptosis of granulosa cells by the PI3K/ Akt/mTOR signaling pathway is ROS-mediated, we added miRNA-146 to determine whether PI3K/Akt/mTOR expression and apoptosis of granulosa cells with the presence of OX-LDL could be suppressed. The results showed that miRNA-146 pretreatment up-regulated the critical mTOR signaling expression in granulosa cells treated with 400 μg/mL OX-LDL (Figure 3A, 3D), which suggested that ROS was activated before the mTOR signaling pathway in OX-LDL-promoted poptosis of granulosa cells. We also found that PI3K and Akt expression of granulosa cells decreased under 400 μg/mL OX-LDL and increased under miRNA-146 with or without OX-LDL (Figure 3A, B and C). Moreover, the poptosis of granulosa cells related genes caspase 3 decreased when treating the poptosis of granulosa cells with miRNA-146 (Figure 2A, 2D). The protein expression of Beclin1 was reduced (Figure 2A, 2B) in OX-LDL-treated granulosa cells. The ratio of LC3II:LC3I was also decreased (Figure 2A and 2C) in OX-LDL- treated granulosa cells. The protein expression of Beclin1 and the ratio of LC3II:LC3I were increased with miRNA-146 in OX-LDL-treated granulosa cells. Takern together, these results dermonstrated that OX-LDL could promote insufficient autophagy and poptosis of granulosa cells in vitro through the ROS-mediated PI3K/Akt/mTOR pathway. Western blotting demonstrated that OX-LDL could not only regulate PI3K/Akt/mTOR signaling pathway, but also stimulate the expression of caspase3. In addition, we proved that miRNA-146 pretreatment could inhibit OX-LDL promoted ROS-mediated PI3K/Akt/ mTOR expression. miRNA-146 also suppressed the expression of caspase 3.
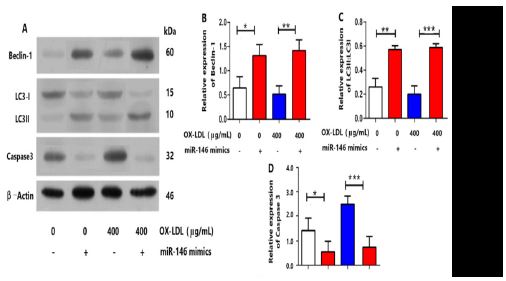
(A) The protein expression levels of mTOR, PI3K and Akt were suppressed under 400 μg/mL OX-LDL and promoted by miRNA-146 mimics with 400 μg/mL OX-LDL or without treatment as examined by Western blot (n = 5 per group). (B,C,D) granulosa cells were pretreated with 100 nmol/L miRNA-146 mimics for 1 h before being cultured with or without 400 μg/mL OX-LDL.
Discussion
POI is a oxidative stress induced chronic inflammatory reaction of the ovary with functional decline. Although ROS is a natural by-product of aerobic metabolism, but unlimited ROS can lead to oxidative stress and cell damage [9]. In the process of reproduction, the increase of energy and nutrient demand is related to the acceleration of metabolic rate and thus the cumulative ROS production. Growing evidence implicates significant contributions of oxidative stress to initiate granulosa cell death through programmed cell death during follicular atresia [13], which may give rise to certain anovulatory disorders, such as POI. In particular, autophagic signals are recognized only in granulosa cells within antral follicles. Importantly, observations of human ovaries reveal a close correlation between OX-LDL-induced autophagy and granulosa cells death [14], these findings indicate a distinctrole of autophagy in regulating granulosa cells viability. Granulosa cells (GCs) are somatic cells originating from the sex cord, which are the crucial type of cells in the ovary. As the major source of oestradiol and oestrogens in response to FSH, GCs have critical roles in oocyte maturation via oocyte–granulose cell interaction and in normal folliculogenesis. In other words, dysfunction of GCs usually leads to pathological folliculogenesis, including premature ovarian failure (POF), polycystic ovary syndrome (PCOS) and ovarian GC tumours (GCT). In addition, ROS levels in specific tissues lead to different processes of cell death. For example, increased ROS levels leads to granulosa cell apoptosis [15]. In humans, granulosa cells exposure to OX-LDL causes autophagy, which further leads to apoptosis and death. According to the study, POI has excess OX-LDL resulting in autophagy dysregulation, apoptosis and cell death of ovarian granulosa cells. Regression autophagy has been found to be rich in oxidized low density lipoprotein (lectin like) receptor 1 (OLR1) in human ovaries [16]. Thus, the OX-LDL-OLR-ROS pathway induces autophagy in granulosa cells [17]. It is reported that autophagy is involved in regulating the apoptosis of granulosa cells to accelerate follicular atresia [18,19]. However, limited information is known about the role of OX- LDL direct induction of insufficient autophagy and apoptosis of ovarian granulosa cells in POI. In this study, we demonstrated for the first time that OX-LDL induced autophagy dysregulation in mice due to elevated OX-LDL in vivo due to regulation of mTOR expression. In addition, we found that OX-LDL directly promotes insufficient autophagy and apoptosis of granulosa cells in vitro through activation of the ROS-mediated PI3K/Akt/mTOR Pathway. These findings have important implications for further understanding the mechanism by which OX-LDL contributes to autophagy dysregulation and granulosa cells apoptosis in POI.
Ovarian dysfunction has the common pathophysiological mechanisms to oxidative stress and chronic inflammation, including OX-LDL and ROS levels [20]. In this study, it showed that OX-LDL can significantly reduce the viability of granulosa cells in a dose-dependent manner. OX-LDL has been proved to be a marker of oxidative stress and a risk factor of various chronic inflammatory diseases. In the current study, we firstly showed that autophagy dysregulation and reduced autophagy level in granulosa cells of POI models. Under normal conditions, sufficient autophagy ensures the appropriate level of mTOR protein to facilitate the differentiation of granulosa cells, whereas with insufficient autophagy, the excessive accumulation of ROS inhibits the transcriptional activation of mTOR and PI3K/Akt, which leads to defective differentiation and the decrease of growth viability of granulosa cells. Therefore, the reduced LC3-II:LC3-I ratio indicates that autophagy defects [21] are down-regulated in autophagy-related genes in the current study, including mTOR and PI3K/Akt. The lower LC3-II: LC3-I ratio and Beclin-1 level indicate that it is the insufficient autophagy in granulosa cells of POI models, which indicates the potential contribution of autophagy disorder to POI pathology. In addition, this study showed that ovarian granulosa cell autophagy deficiency and apoptosis coexist under OX-LDL exposure.
mTOR belongs to the PI3K-related kinase (PIKK) family and is associated with ROS [22]. PI3K triggers Akt to activate mTOR. In most cases, mTOR is controlled by a kinase cascade involving PI3K and Akt [23]. The stimulation of PI3K/Akt dependent cascade may reduce the apoptosis and death of ovarian granulosa cells. Previous investigator indicated that PI3K/Akt/ mTOR pathway is activated by melatonin to inhibit autophagy protection against POI [24]. Yan et al. also demonstrated that the PI3K/Akt/mTOR signal pathway can protect rat ovarian granulosa cells from oxidative stress and apoptosis [25]. Chen et al. reported that acupuncture regulates autophagy of ovarian granulosa cells in patients with ovulation disorders by inhibiting PI3K/AKT/ mTOR pathway with LncMEG3 [26]. Regulation of PI3K/AKT/mTOR pathway can reduce excessive autophagy of ovarian granulosa cells [27]. Herein, our data implied that ROS increases the down-regulation expression of PI3K/Akt/mTOR in granulosa cells in OX-LDL- induced in vivo and in vitro. miRNA-146 can inhibit the expression of ROS, which can rescue the phosphorylation of the above proteins. Moreover, the expressions of the phosphorylated and non-phosphorylated PI3K/Akt/mTOR signalling pathways in the miRNA-146 pretreatment group showed significant alterations compared with those in the control groups, which indicated that ROS acted as an upstream event of PI3K, Akt and mTOR [28]. The analytical results suggested that OX-LDL promotes insufficient autophagy and apoptosis of ovarian granulosa cells through ROS-mediated PI3K/AKT/mTOR signalling pathway. However, our study has some limitations. Although it is to study the role and mechanism of OX-LDL exposure in the occurrence and development of POI. The regulation of OX-LDL on ovarian granulosa cells is not only PI3K/AKT/mTOR pathway. In addition, we only studied the effect of miRNA-146 on the abnormality of OX-LDL activating ROS-mediated PI3K/AKT/mTOR pathway in ovarian granulosa cells of POI mouse model, but it is not clear that miRNA-146 regulates OX-LDL activating ROS-mediated PI3K/AKT/mTOR pathway affects the mechanism of insufficient autophagy in granulosa cells. Therefore, we are planning to carry out a clinical study to expand the sample to understand the mechanism of miRNA-146 inhibiting OX-LDL activating ROS- mediated PI3K/AKT/mTOR signal pathway to ameliorate ovarian function and reduce or inhibit granulocyte autophagy dysregulation and apoptosis.
Conclusion
To our knowledge, this study is the first to demonstrate that OX-LDL promotes insufficient autophagy, and apoptosis of ovarian granulosa cells through ROS-mediated PI3K/AKT/mTOR signalling pathway. In addition, the current study showed that under OX-LDL exposure, ovarian granulosa cells insufficient autophagy and apoptosis coexist in the POI mouse models. Besides, OX-LDL could directly induce ovarian granulosa cells apoptosis, and miRNA-146 could inhibit it. Overall, the present findings provide detailed information for understanding the exact mechanism of insufficient autophagy and apoptosis coexist in granulosa cells under OX-LDL.
Declarations
Disclosure statement : The authors declared that they do not have any commercial or associative interest that represents a conflict of interest in connection with the work submitted.
Funding : This work was supported by Major Project of Regional collaborative Fund of Foundational and Basic Research for Application of Guangdong Prevince (No. 2020B1515120009).
References
- Costa GPO, Ferreira-Filho ES, Simoes RDS, Soares-Junior JM, Baracat EC, et al. Impact of hormone therapy on the bone density of women with prematureovarian insufficiency: A systematic review. Maturitas. 2023; 167: 105-112.
- Zhang X, Lu Y, Wu S, Zhao X, Li S, et al. Estimates of global research productivity in primary ovarian insufficiency from 2000 to 2021: Bibliometric analysis. Front Endocrinol (Lausanne). 2022; 13: 959905.
- McGlacken-Byrne SM, Conway GS. Premature ovarian insufficiency. Best Pract Res Clin Obstet Gynaecol. 2022; 81: 98-110.
- An R, Wang X, Yang L, Zhang J, Wang N, et al. Polystyrene microplastics cause granulosa cells apoptosis and fibrosis in ovary through oxidative stress in rats.Toxicology. 2021; 449: 152665.
- Fengping He, Yanhui Liu, Li Tang, Qiulin Ma , Yongmei Zhang,Peiqing He , and Chuanyin Xiong. MicroRNA-146 attenuates lipopolysaccharide induced ovarian dysfunction by inhibiting the TLR4/NF- κB signaling pathway. Bioengineered. 2022; 13: 1161-116232.
- Chen C, Li S, Hu C, Cao W, Fu Q, et al. Protective Effects of Puerarin on Premature Ovarian Failure via Regulation of Wnt/β-catenin Signaling Pathway and Oxidative Stress. Reprod Sci. 2022;28:982-990.
- Jing Zhou, Jun Zhou, Liu-Jian-Xiong Wu,Yi-Yang Li, et al. CircRNA circUSP36 impairs the stability of NEDD4L mRNA through recruiting PTBP1 to enhance ULK1-mediated autophagic granulosa cell death. J Reprod Immunol. 2022; 153:103681.
- Liu X, Lin X, Zhang S, Guo C, Li J, et al. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging (Albany NY). 2018; 10: 2016-2036.
- Prasad S, Tiwari M, Pandey AN, Shrivastav TG, Chaube SK. Impact of stress on oocyte quality and reproductive outcome. J Biomed Sci. 2016; 23: 36.
- Sul OJ, Ra SW. Quercetin Prevents LPS-Induced Oxidative Stress and Inflammation by Modulating NOX2/ROS/NF-kB in Lung Epithelial Cells. Molecules. 2021; 26: 6949.
- Katharina Spanel-Borowski, Marcin Nowicki, Juergen Borlak, Tom, et al. Endoplasmic reticulum-derived multilamellar bodies in oocytes of mouse follicle cultures under oxidized low-density lipoprotein treatment. Cells Tissues Organs. 2013; 197: 77-88.
- Ying Liu, Jianjun Zhai, Jing Chen, Xiaomin Wang, Tingru Wen. PGC-1α protects against oxidized low-density lipoprotein and luteinizing hormone-induced granulosa cells injury through ROS-p38 pathway. Hum Cell. 2019; 32: 285-296.
- Wang L, Tang J, Wang L, Tan F, Song H, et al. Oxidative stress in oocyte aging and female reproduction. J Cell Physiol. 2021; 236: 7966-7983.
- Zhou J, Zhou J, Wu LJ, Li YY, Li MQ, et al. CircRNA circUSP36 impairs the stability of NEDD4L mRNA through recruiting PTBP1 to enhance ULK1- mediated autophagic granulosa cell death. J Reprod Immunol. 2022; 153: 103681.
- Prasad S, Tiwari M, Pandey AN, Shrivastav TG, Chaube SK. Impact of stress on oocyte quality and reproductive outcome. J Biomed Sci. 2016; 23: 36.
- Sahin SB, Nalkiran I, Ayaz T, Irfan Guzel A, et al. Genetic variations in OLR1 gene associated with PCOS and atherosclerotic risk factors.J Investig Med. 2023; 71: 113-123.
- Weitzel JM, Vernunft A, Krüger B, Plinski C, Viergutz T. LOX-1 regulates estrogenesis via intracellular calcium release from bovine granulosa cells. Cytometry A. 2014; 85: 88- 93.
- Sahin SB, Nalkiran I, Ayaz T, Irfan Guzel A, Eldes T, Calapoglu T, Sevim Nalkiran H. Genetic variations in OLR1 gene associated with PCOS and atherosclerotic risk factors. J Investig Med. 2023; 71: 113-123.
- Shao T, Ke H, Liu R, Xu L, Han S, et al. Autophagy regulates differentiation of ovarian granulosa cells through degradation of WT1. Autophagy. 2022; 18: 1864-1878.
- Tesarik J, Galán-Lázaro M, Mendoza-Tesarik R. Ovarian Aging: Molecular Mechanisms and Medical Management. Int J Mol Sci. 2021; 22: 1371. .
- Schaaf MB, Keulers TG, Vooijs MA, et al. LC3/GABARAP family proteins: autophagy-(un)related functions. FASEB J. 2016; 30: 3961-3978.
- Sanchez-Martin P, Komatsu M. p62/SQSTM1 - steering the cell through health and disease. J Cell Sci. 2018; 131: jcs222836.
- Liu Y, Zhu X, Wu C, Lang Y, Zhao W, et al. Melatonin protects against ovarian adamage by inhibiting autophagy in granulosa cells in rats. Clinics (Sao Paulo). 2022; 77: 100119.
- Liu Y, Fang Y, Wei J, Zhang C, Wu D, et al. Melatonin Protects against Primary Ovarian Insufficiency by Activating the PI3K/Akt/mTOR Pathway and Inhibiting Autophagy. Ann Clin Lab Sci. 2022; 52: 895-903.
- Yan J , Deng D, Wu Y, Wu K, Qu J, et al. Catalpol protects rat ovarian granulosa cells against oxidative stress and apoptosis through modulating the PI3K/Akt/Mtor signaling pathway. Catapol Biosci Rep. 2020; 40: BSR20194032.
- Xiaohong Chen, Hongliang Tang, Yingye Liang, Pingting Wu, Liuhong Xie, et al. Acupuncture regulates the autophagy of ovarian granulosa cells in polycystic ovarian syndrome ovulation disorder by inhibiting the PI3K/AKT/mTOR pathway through LncMEG3. Biomed Pharmacother. 2021; 144: 112288.
- Xu B, Dai W, Liu L, Han H, Zhang J, et al. Metformin ameliorates polycystic ovary syndrome in a rat model by decreasing excessive autophagy in ovarian granulosa cells via the PI3K/AKT/mTOR pathway. Endocr J. 2022; 69: 863-875.
- Kma L, Baruah TJ. The interplay of ROS and the PI3K/Akt pathway in autophagy regulation.Biotechnol Appl Biochem. 2022; 69: 248-264.